

Subscribe to RSS
DOI: 10.1055/s-0044-1790517
Sheep Head Cadaveric Model for the Transmeatal Extensions of the Retrosigmoid Approach
Authors

Abstract
The transmeatal extension of the retrosigmoid approach is an important procedure used in the treatment of various pathologies affecting the posterior fossa, petroclival region, and jugular foramen. Mastering this technique requires a high level of manual skill, particularly in temporal bone drilling. The objective of this study was to describe an easily accessible and cost-effective model of the transmeatal extension of the retrosigmoid approach using cadaveric sheep heads. Five cadaveric sheep heads, fixed in alcohol and formalin with intravascular-colored silicone injection, were prepared for this study. Two heads (four sides) were designated for illustrative anatomical specimens, while three heads (six sides) were used for surgical simulation. Additionally, one head was used to prepare and dissect a dry skull. All critical steps of the transmeatal approach, including both supra- and inframeatal extensions, were successfully replicated on the model. A comparative anatomical analysis was conducted, focusing on the technical nuances of the model. The cadaveric sheep head serves as an effective model for the retrosigmoid approach with transmeatal extensions, primarily for training manual haptic skills. While the sheep model cannot precisely replicate human anatomy, it still offers valuable training opportunities for neurosurgeons, particularly when human cadaveric specimens are unavailable.
Keywords
training model - cadaveric model - sheep head - temporal bone dissection - retrosigmoid approach - retrosigmoid approach extensions - suprameatal - transmeatal - inframeatal - suprajugular approachIntroduction
Temporal bone serves as a gateway to diverse surgical corridors, offering access to lesions within the posterior and middle fossa, including those that involve both areas.[1] [2] The part of the temporal bone that poses the most significant challenge in surgery is the petrous part. Being pyramid-shaped, the petrous part encases important neurovascular structures. Violation of these structures can lead to serious morbidity, impacting the patient's quality of life. Damage to the vestibulocochlear nerve complex and inner ear structures, such as the labyrinth or endolymphatic sac, can result in hearing loss, injury to the facial nerve can lead to facial nerve palsy, while damage to the petrous carotid artery and jugular vein can cause life-threatening bleeding. Manipulation of the sigmoid sinus may result in postoperative thrombosis, and even manipulation of the air cells of the temporal bone can lead to postoperative cerebrospinal fluid leakage. The matter is further complicated by the fact that all these structures are tightly enclosed within the petrous part of the temporal bone. The pathologies of the cerebellopontine angle (CPA) often affect the temporal bone. Vestibular shwannomas, one of the most common pathologies of the CPA, invade the internal acoustic canal. Chordomas and chondromas, originating in the bone, encase the neurovascular structures within their parenchyma. Increasing the radicality of tumor resection offers higher chances to improve postoperative outcomes, reduce morbidity, mortality, and recurrence rates.[1] However, on the other hand, it increases iatrogenic complications. The lack of mastery of manual skills by the surgeon and fear of causing iatrogenic harm often lead neurosurgeons to avoid the radicality in surgical treatment of these pathologies.
Mastering temporal bone dissection depends significantly on both manual skills for bone dissection and a firm understanding of anatomy, along with the ability to three-dimensional (3D) interpretation of preoperative neuroimaging studies. Hence, the model able to encompass both these skills may be considered as the best option. Though animal cadaveric models are commonly used in training for skull base and ear, nose, and throat surgeons in some institutions,[3] [4] [5] [6] [7] [8] they can offer only the realistic haptic feedback experience, lacking in aspects of anatomical similarity with human. But at the same time the skill of orientation relying on analytical orientation and not visual also may be trained on animal models.[9] [10] The human cadaveric model, essentially includes both modalities of surgical training, and deserves to be regarded as the best model.[11] [12] The human cadaveric material is not always easily accessible because of costs of cultural specificities of different countries. The increasing popularity of digital technologies and 3D printing has led to the emergence of synthetic 3D-printed models.[13] [14] [15] These models offer advantages such as lower costs and the absence of ethical issues. While they generally simulate human anatomy, they cannot replicate the tactile experience of exercising on biological tissues.
The goal of this study was not to oppose the animal model to human cadaveric, synthetic, or virtual models, but rather to highlight the undeniable advantages of this model, which may play a beneficial role in developing and refining the skill for bone dissection necessary for internal acoustic meatotomy procedure.
Materials and Methods
Six cadaveric sheep heads were used for this study. The material for this study was obtained from local slaughterhouse. Sheep heads were delivered frozen. The special statement to be made is that no animals were sacrificed specifically for the purpose of this study. To enhance the study of anatomy and facilitate the recognition of intracranial vascular structures, portions of the heads were fixed, and colored silicone was injected into the carotid arteries and jugular veins.
For this study four sides of two sheep cadaveric heads were dissected in order to prepare anatomical specimens to perform comparative analysis between human and sheep anatomy. Six sides of three heads in order to simulate the retrosigmoid extended approach.
Microsurgical stage was performed using the laboratory microscope (Leica M 320 T, Germany) with magnification (×4– × 12). Craniotomy and microdrilling were performed using drill (Marathon, China) 120 W RS 0-30,000 rpm/min, photos were made using professional digital photo camera (Nikon, Thailand).
Techniques
Five cadaveric sheep heads, purchased in the local slaughterhouse, were used for the study. The fixation and intravascular silicone injection were performed guided by published papers.[16] [17] Sheep heads were washed in warm running water. Common carotid arteries and jugular veins were dissected on both sides, cannulated with plastic nasogastric catheters 8 to 12 Fr, and ligated with silk threads to vessel wall. Under moderate pressure vessels were washed with warm water, to remove blood and clots from the vessels.
The fixation of the specimens was achieved by perfusion of both carotid arteries and jugular veins with a mixture of 10% alcohol and 10% formalin in a 1/1 ratio for 15 minutes, followed by immersion in a 10% formalin solution. Intravascular injection of the alcohol-formalin mixture was repeatedly performed every 8 hours for 3 days. After 3 days of exposition colored silicone (Redelease, Brazil) was injected, red to the arteries and blue to the veins. Heads were exposed for 5 days more in 10% formalin solution.
Prior to dissection the heads were soaked in warm water for several hours to reduce the formalin odor. Heads were stabilized with the help of Mayfield headholder (Makom, Brazil) ([Fig. 1]). For better demonstration of all the steps of the retrosigmoid approach all layers of the scalp and nuchal area were preserved, wool along the incision lines may be shaved, however, this step is not necessary.

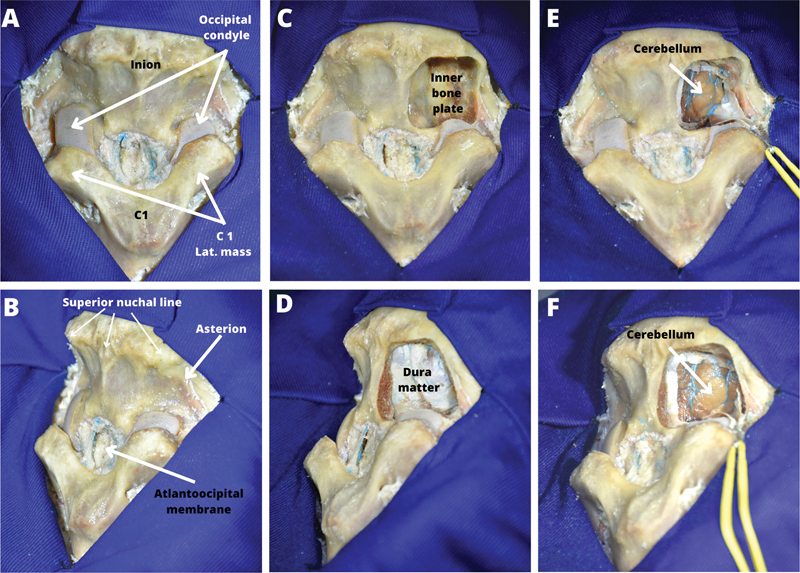
The nuchal muscles may be dissected layer by layer, which may serve as a good exercise by itself ([Fig. 2]). Another way is that after the dissection of the superficial layer of the nuchal muscles, the paravertebral deep muscle complex may be cut on midline along the nuchal ligament and separated from the occipital bone and the first cervical vertebra ([Fig. 3]). The layer by layer dissection of the soft tissues of occipito-nuchal area was performed using fish hooks retraction. The burr hole was placed on the point of asterion, using anatomical landmarks with comparative references with human anatomy ([Fig. 1]). Taking into account the more intimate adherence of the inner bone plate to the posterior fossa dura, and in order to preserve the dura intact, the craniotomy window was formed in an osteoclastic manner—widening the burr hole using a high-speed drill, or biting with rongeurs. After performing the craniotomy, dura was incised and the flap was retracted laterally ([Fig. 3]). In alcohol-fixed specimens, the preserved elasticity of the brain allows appropriate exposure of the CPA by moderate retraction of the cerebellar hemisphere. If the stiffness of the formalin-fixed brain does not allow sufficient exposure through retraction, the hemisphere may need to be resected.

Results
During the study two principal goals were established: the comparative anatomical analysis and the stepwise simulation of the real surgery.
Comparative Anatomy
To create a technical guide for the exercise and highlight the differences between the exercise conditions and those of actual surgery or human cadavers, special attention was given to the description of ovine anatomy.
During the study, we conducted a detailed descriptive analysis of the CPA anatomy, with particular attention to the internal acoustic meatus in sheep ([Fig. 4]) as well as the broader CPA anatomy ([Figs. 5] and [6]). The ovine anatomy, being at the first glance quite different from the human, still follows the same principles that underly and govern the relations between the anatomical structures in both species. The anatomical analysis was structured as a distinct comparison between osseous, neural, and vascular anatomy, and presented in photos and diagrams ([Figs. 7] [8] [9]).






Osseous Anatomy
The initially noticeable difference in osseous anatomy is the size of the skull. Due to the relatively smaller size of the posterior fossa in sheep compared to humans, a relatively larger craniotomy is required to adequately expose the CPA on a sheep model ([Figs. 7] [8] [9]). The internal acoustic meatus represents a prominent structure that bulges to the CPA in a tube-like fashion ([Figs. 4] and [8]).
The interior acoustic meatus is shallower than in humans, with a depth of no more than 2 to 3 mm. The fundus of the internal acoustic meatus is also divided into four compartments, the transverse crest is well-defined bulky osseous wall that divides the meatus into upper and lower semicanals in the horizontal plane ([Fig. 4B]). The Bill's bar is also well identified. The facial nerve canal in general follows a similar route to that in the human petrous bone. The facial canal starts at the upper inner quarter of the internal acoustic meatus, heads forward, splits into the canal of the greater petrosal nerve, then curves backwards. It passes under the lateral semicircular canal and enters the tympanic cavity ([Fig. 9]). The arcuate eminence, a bony protuberance with the superior semicircular canal underneath it, is a prominent feature on the superior surface of the petrous bone. Just like in humans, it serves as a reliable landmark for identifying the superior semicircular canal and the common crus of the superior and posterior canals ([Fig. 9]).
Vascular Anatomy
The internal carotid is not present as a single arterial trunk, but as s rete carotideo—several efferent arterial vessels creating arterial plexus, also called as rete mirabile epidurale rostrale which lies inside the cavernous sinus instead of intracavernous carotid. So, the carotid canal is not opening on the extracranial skull base of the petrous bone but lies in the laceral fissure—the cleft-like space inside the petroclival suture ([Figs. 4] and [7]). The anterior inferior cerebellar artery (AICA) is not present as a well-organized, single-trunk tortuous vessel with the typical loop shape. The sigmoid sinus and the jugular bulb are not present in ovine anatomy. The main venous collector of intracranial venous blood is the maxillary vein. The maxillary vein receives the major flow of the venous blood from the transverse sinus. However, there is a venous trunk passing inside the thickness of the mastoid portion of the temporal bone in the same direction as a sigmoid sinus in human and joining the internal jugular vein through the caudal division of the jugular foramen, which apparently corresponds to the venous part of the jugular foramen.
Cranial Nerves
Generally speaking, the cranial nerves of the vestibulocochlear complex are proportionally thicker and shorter than in human. The cranial nerves of the CPA follow distinct trajectories, allowing for their visual recognition. The direction of the nerves is the axis from the brainstem origin of the nerve and the cranial foramina—porus trigeminus, internal acoustic meatus, and jugular foramen. In human anatomy the plane of the axis of the trigeminal nerve and the axis of vestibular facial complex and the plane of the caudal nerves and vestibular facial nerves cross in open angle while in sheep they almost lie in a single plane ([Fig. 9]).
Vestibular Labyrinth and the Semicircular Canals
In general features, such as the orientation, the angle between the planes of the semicircular canals and the correspondence to the other structures of the temporal bone is surprisingly similar to the human labyrinth ([Figs. 7] and [9]).
Surgery Imitation
During the study the imitation of the retrosigmoid extended approach was successfully performed. The focus was made on the simulation of the key sequential steps for each type of the transmeatal extensions.
Numerous variants of the retrosigmoid approach's extensions involving dissections of the internal acoustic meatus were described in the literature nowadays.[18] [19] [20] [21] [22] [23] [24] [25] [26] [27] [28] [29] [30] [31] [32] [33] [34] [35] [36] [37] [38] [39] [40] [41] [42] For the sake of simplification, the extensions of retrosigmoid approach may be roughly divided in two groups: above the internal acoustic meatus and below the acoustic meatus; however, both of those groups require wide exposure of the fundus of the internal acoustic meatus.
The perimeatal extensions of the retrosigmoid approach were organized into two groups—supra- and inframeatal extensions. The procedures were subdivided into principal stages, each of them supposed to be reproduced on the sheep head model. As an additional step apicectomy was mentioned. The imitation of the extensions of the retrosigmoid approach was successfully performed ([Fig. 10], [Videos 1] and [2]).
Video 1 Retrosigmoid suprameatal and transapical approach.

Both of the groups of extensions of retrosigmoid approach were imitated, taking into account anatomical differences between ovine and human temporal bone anatomy, following certain steps.
Transmeatal Extension ([Videos 1] and [2])
-
(1) Cut the dura over internal auditory canal (IAC) (identify endolymphatic sac as a landmark for trajectory selection; Tubingen line between jugular foramen and IAC); endolymphatic duct leads to the crus of labyrinth—confluence of superior and posterior semicircular canals—as in human ([Fig. 9]).
-
(2) Drilling of the posterior wall of the internal acoustic meatus until the appearance of the transverse crest ([Fig. 5]).
-
(3) Dissection of the facial nerve from vestibular nerves ([Fig. 9]).
Suprameatal Extension (Retrosigmoid Intradural Suprameatal Approach) ([Video 1])
-
(1) Cut the dura over IAC, over suprameatal tubercle (landmarks to identify superior semicircular) ([Fig. 10]).
-
(2) Drilling the roof of the IAC and tentorial surface of the petrous apex ([Fig. 10F]′).
-
(3) Mobilize the lateral wall of Meckel's cave, dissect the back wall of the cavernous sinus, transect the tentorial edge, and dissect trochlear nerve in case of reversed Kawase extension ([Fig. 10E]′, F′).
Inframeatal and Suprajugular Extension ([Video 2])
Video 2 Retrosigmoid transmeatal suprajugular approach.
-
(1) Cut the dura posteriorly to IAC and jugular foramen, perpendicular to Tubingen line (landmarks to identify cochlea) ([Fig. 10C]).
-
(2) Drilling the posteroinferior wall of the internal acoustic meatus ([Fig. 10D′]).
-
(3) Incision of intra-acoustic and intrajugular dura ([Fig. 10E, F]).
-
(4) Unroofing of the jugular foramen ([Fig. 10E, F]).
The terms of the surgical simulation in the presented model were defined by three points: (1) technical skills of handling arachnoid dissection of CPA nerves; (2) identification and recognition of CPA nerves anatomy; and (3) intradural drilling of the medial wall of the temporal bone.
Discussion
Importance of the Extensions of the Retrosigmoid Approach
The retrosigmoid approach is a workhorse approach to posterior fossa lesions. Performed with extensions based on intradural temporal bone drilling, it significantly widens microsurgical routes to all levels of the posterior fossa as well as the posterior wall of the cavernous sinus and petroclival region. The selection of one of the extensions depends on the objective of the approach.[18] [19] [20] [21] [22] The transmeatal approach, that requires the opening of the posterior wall of the internal acoustic meatus, is utilized in most cases of vestibular schwannoma with intrameatal growth.[23] [24] [25] [26] [27] [28] [29] [30] [31] For petroclival tumors, or those located anterior to the vestibulocochlear nerve complex, the retrosigmoid intradural suprameatal approach (RISA) is employed, necessitating the resection of the upper wall of the internal acoustic meatus.[32] [33] [34] [35] [36] In case of tumors of the Meckel's cave invading the medial cranial fossa, posterior intradural petrous apexectomy provides access to the middle cranial fossa from the posterior fossa, by resecting the petrous apex.[34] [36] Inframeatal extension serves as an approach to tumors of jugular foramen region and requires drilling the jugular tubercle, intrajugular process, and medial side of the pyramid apex.[37] [38] [39] [40] [41] [42] Possibility of sparing facial nerve in vestibular schwannoma surgery by opening the back wall of the internal acoustic meatus makes transmeatal extension of the retrosigmoid approach an excellent alternative to transpetrosal translabyrinthine approach and anterior transpetrosal approach.[43] [44] [45] Being less invasive, performing retrosigmoid approach may decrease intraoperative time, being at the same time no less effective.
CPA is a complex anatomical region containing three neurovascular complexes of cranial nerves and cerebellar arteries. Drilling in a narrow space with neighborhood of cranial nerves and brainstem requires perfect control of a drill handpiece and the tip of rotating burr, tactile and spatial perception of the interfaces between bones to be drilled.
Drilling the posterior wall of the internal acoustic canal has technical challenges and risks, such as damage of inner ear structures such as the labyrinth, cochlea, facial nerve, and endolymphatic duct. Numerous anatomic studies were performed to make temporal bone drilling safer and surgeries more radical.[45] [46] [47] [48] [49] [50]
One of the main obstacles on the way of introduction of such techniques to routine practice of the majority of institutions is a lack of hands-on training on human cadaveric models because of high costs and legal and cultural issues in a row of countries.
The principal point of using human cadaveric models is the possibility to get familiar with normal human anatomy and this point is the only advantage in which animal models fails to be more effective.
Advantages and Limitations
First of all, we need to clarify that the choice of the sheep head was not based on any comparative analysis with other animal models. We simply used the model that was most readily available to us and are presenting our experience with it as an additional low-cost biological model option.
Systematizing the differences and similarities that were listed in the section “Results,” and speaking about the anatomical comparisons in general, we need to make specific assertions. The criteria for any comparison may be divided into two groups: quantitative and qualitative. Quantitative characteristics of anatomical structures include size, volume, shape, and angles between their axes. On the other hand, qualitative characteristics encompass the presence or absence of specific structures, their numbers, entry and exit points of nerves and vessels in bony openings, and their topographical relationships with neighboring structures.
Quantitative comparison: (1) internal acoustic meatus is more shallow and proportionally wider than one in human—creates different angle of attack during posterior wall drilling. (2) Facial nerve and the nerves of vestibulocochlear complex are relatively thicker and much shorter—makes them more vulnerable to avulsion from the brainstem, thus requiring more delicate manipulations.
Qualitative comparison: (1) the jugular foramen does not contain the jugular vein—that devoid the model of the important skill of managing the jugular bulb in case of inframeatal extension. (2) The carotid canal is located more parallel to the clival axis, contains rete carotideo, but not a single arterial trunk.
In cases of normal human anatomy, comparisons between human specimens are predominantly quantitative. This means that during cadaveric dissection, the trainee sees more or less the same anatomical picture, with insignificant differences. The same applies to anatomical atlases. Neurosurgeons become accustomed to the subtle variations in normal anatomy, causing quantitative characteristics to become qualitative in their perception. The surgeon learns to recognize the structures by their typical size, shape, and location and not by their relationships to other important structures. For instance, the facial nerve may have a different diameter, shape, and pass in different directions, but it will never originate from anywhere else but pontomedullary junction, and end in the upper inner quadrant of the internal acoustic meatus. Significant changes in size, shape, or location of the customary structure make it unrecognizable, and may confuse the surgeon. This challenge is exactly what occurs in case of pathologically distorted anatomy. The lesion leads to qualitative changes, such as displacement, destruction, or deformation of adjacent structures due to tumor growth or invasion. This may complicate the process of orientation, and hinder correct decision making.
Training on an animal model challenges the trainee in the same manner as pathologically altered anatomy. The potential for confusion and misunderstanding during exercises involving anatomical discrepancies between human and sheep anatomy mirrors the challenges faced in actual surgery due to anatomical variations caused by pathology or normal anatomical variants. Despite differences in the anatomical locations of crucial structures, their order and spatial relationships remain consistent with those in humans. Analyzing the animal's anatomy prepares a trainee to resolve these practical challenges and makes the thinking flexible and analytical.
The interfaces between tissues with different densities, elasticities, and structures with complex geometrical shapes cannot be imitated in synthetic or virtual models better than in biological cadaveric material. Therefore, using real cadaveric material, even though animal, in training offers superior tactile feedback to trainees. This allows to expect higher efficiency of the training for the identification and dissection of structures such as the dural folds and sheaths, cranial nerves of the CPA, internal acoustic meatus anatomy with its compartments, and proximity to the labyrinth and cochlea.
The low costs of the animal cadaveric training model also makes it accessible, in low-income countries, and raises fewer ethical and cultural concerns compared to training with human cadavers.
The tactile experience received on the animal model remains similar to human cadaver, and the visually different anatomy of other species requires deep analysis of the general anatomical principal to which both human and animal obey. This analytical skill is very useful when the surgeon who studied human anatomy on normal specimen meets the anatomy in the surgical room. Being sometimes variative or distorted by pathology, even human anatomy in actual surgery may be hardly recognizable if one relies on visual recognition but not on systemic analytical judgment of the anatomy. As have been told above, the same laws govern the arrangement and relationships between the vital structures as well as in human and animal also as in normal and pathologically distorted anatomy in the human. The skill of structuralized anatomical analysis is an extremely useful skill to be confident in actual procedure.
Conclusion
The presented cadaveric sheep head model for the extended retrosigmoid approach may serve as a cost-effective alternative to human cadaveric temporal bone, in regard of polishing of manual skill and gaining tactile experience in acoustic meatotomy and orientation in CPA anatomy. Although the model has poor anatomical similarity to humans in terms of quantitative comparison of neurovascular and osseous structures, hence cannot be considered a tool for learning human anatomy. However, qualitative similarity between human and sheep makes it a valuable tool for training the capacity for deep structured anatomical analysis. It is essential to emphasize that this study does not aim to oppose the sheep model to the human cadaveric model or to assert the similarity between human and ovine anatomy. Instead, the anatomical analysis presented in this study provides a concise guide for trainees, focusing on the practical aspects of surgical training rather than anatomical similarity. Sheep head model serves as an appropriate tool in mastering intradural drilling of the inner side of the petrous bone. Such technical difficulties as drilling in deep narrow space, orientation in CPA using cranial nerves osseous landmarks, and bone drilling without injuring of neurovascular structures may be successfully simulated on the presented model.
Conflict of Interest
None declared.
-
References
- 1 Basma J, Anagnostopoulos C, Tudose A. et al. History, variations, and extensions of the retrosigmoid approach: anatomical and literature review. J Neurol Surg B Skull Base 2021; 83 (Suppl. 02) e324-e335
- 2 Rodriguez Rubio R, Xie W, Vigo V. et al. Immersive surgical anatomy of the retrosigmoid approach. Cureus 2021; 13 (06) e16068
- 3 Al-Sharshahi ZF, Hoz SS, Alrawi MA, Sabah MA, Albanaa SA, Moscote-Salazar LR. The use of non-living animals as simulation models for cranial neurosurgical procedures: a literature review. Chin Neurosurg J 2020; 6: 24
- 4 Mantokoudis G, Huth ME, Weisstanner C. et al. Lamb temporal bone as a surgical training model of round window cochlear implant electrode insertion. Otol Neurotol 2016; 37 (01) 52-56
- 5 Cordero A, del mar Medina M, Alonso A, Labatut T. Stapedectomy in sheep: an animal model for surgical training. Otol Neurotol 2011; 32 (05) 742-747
- 6 Gocer C, Eryilmaz A, Genc U, Dagli M, Karabulut H, Iriz A. An alternative model for stapedectomy training in residency program: sheep cadaver ear. Eur Arch Otorhinolaryngol 2007; 264 (12) 1409-1412
- 7 Hamamcioglu MK, Hicdonmez T, Tiryaki M, Cobanoglu S. A laboratory training model in fresh cadaveric sheep brain for microneurosurgical dissection of cranial nerves in posterior fossa. Br J Neurosurg 2008; 22 (06) 769-771
- 8 Turan Suslu H, Ceylan D, Tatarlı N. et al. Laboratory training in the retrosigmoid approach using cadaveric silicone injected cow brain. Br J Neurosurg 2013; 27 (06) 812-814 Erratum in: Br J Neurosurg. 2014;28(6):823. Bahrı, Yasar [corrected to Bayrı, Yasar]
- 9 Sudhakara Rao M, Chandrasekhara Rao K, Raja Lakshmi C, Satish Chandra T, Murthy PSN. Suitable alternative for human cadaver temporal bone dissection: comparative micro ear anatomy of cattle, pig and sheep with human. Indian J Otolaryngol Head Neck Surg 2019; 71 (04) 422-429
- 10 Mallet C, Cornette R, Guadelli J. Morphometrical distinction between sheep (Ovis aries) and goat (Capra hircus) using the petrosal bone: application on French Protohistoric sites. Int J Osteoarchaeol 2019; 29 (04) 525-537
- 11 Irugu DV, Singh AC, Sikka K, Bhinyaram J, Sharma SC. Establishing a temporal bone laboratory in teaching institutes to train future otorhinolaryngologists and fundamentals of temporal bone laboratory: considerations and requirements. Indian J Otolaryngol Head Neck Surg 2016; 68 (04) 451-455
- 12 Mowry SE, Woodson E, Gubbels S, Carfrae M, Hansen MR. A simple assessment tool for evaluation of cadaveric temporal bone dissection. Laryngoscope 2018; 128 (02) 451-455
- 13 Frithioff A, Frendø M, Pedersen DB, Sørensen MS, Wuyts Andersen SA. 3D-printed models for temporal bone surgical training: a systematic review. Otolaryngol Head Neck Surg 2021; 165 (05) 617-625
- 14 Frithioff A, Frendø M, Weiss K. et al. Effect of 3D-printed models on cadaveric dissection in temporal bone training. OTO Open 2021; 5 (04) X211065012
- 15 Okada DM, de Sousa AM, Huertas RdeA, Suzuki FA. Surgical simulator for temporal bone dissection training. Braz J Otorhinolaryngol 2010; 76 (05) 575-578
- 16 Alvernia JE, Pradilla G, Mertens P, Lanzino G, Tamargo RJ. Latex injection of cadaver heads: technical note. Neurosurgery 2010; 67 (2, suppl operative): 362-367
- 17 Sanan A, Abdel Aziz KM, Janjua RM, van Loveren HR, Keller JT. Colored silicone injection for use in neurosurgical dissections: anatomic technical note. Neurosurgery 1999; 45 (05) 1267-1271 , discussion 1271–1274
- 18 Tatagiba M, Roser F, Schuhmann MU, Ebner FH. Vestibular schwannoma surgery via the retrosigmoid transmeatal approach. Acta Neurochir (Wien) 2014; 156 (02) 421-425 , discussion 425
- 19 Matsushima K, Kohno M, Komune N, Miki K, Matsushima T, Rhoton Jr AL. Suprajugular extension of the retrosigmoid approach: microsurgical anatomy. J Neurosurg 2014; 121 (02) 397-407
- 20 Alimohamadi M, Samii M. Suprajugular extension of the retrosigmoid approach. J Neurosurg 2014; 121 (03) 764-765
- 21 Constanzo F, Gerhardt J, Ramina R. How I do it: retrosigmoid suprajugular approach to the jugular foramen. Acta Neurochir (Wien) 2019; 161 (11) 2271-2274
- 22 Colasanti R, Tailor AR, Zhang J, Ammirati M. Functional petrosectomy via a suboccipital retrosigmoid approach: guidelines and topography. World Neurosurg 2016; 87: 143-154
- 23 Day JD, Kellogg JX, Fukushima T, Giannotta SL. Microsurgical anatomy of the inner surface of the petrous bone: neuroradiological and morphometric analysis as an adjunct to the retrosigmoid transmeatal approach. Neurosurgery 1994; 34 (06) 1003-1008
- 24 Scerrati A, Lee JS, Zhang J, Ammirati M. Exposing the fundus of the internal acoustic meatus without entering the labyrinth using a retrosigmoid approach: is it possible?. World Neurosurg 2016; 91: 357-364
- 25 Scerrati A, Lee JS, Zhang J, Ammirati M. Microsurgical anatomy of the internal acoustic meatus as seen using the retrosigmoid approach. Otol Neurotol 2016; 37 (05) 568-573
- 26 Cueva RA, Chole RA. Maximizing exposure of the internal auditory canal via the retrosigmoid approach: an anatomical, radiological, and surgical study. Otol Neurotol 2018; 39 (07) 916-921
- 27 Miller RS, Pensak ML. An anatomic and radiologic evaluation of access to the lateral internal auditory canal via the retrosigmoid approach and description of an internal labyrinthectomy. Otol Neurotol 2006; 27 (05) 697-704
- 28 Savardekar A, Nagata T, Kiatsoontorn K. et al. Preservation of labyrinthine structures while drilling the posterior wall of the internal auditory canal in surgery of vestibular schwannomas via the retrosigmoid suboccipital approach. World Neurosurg 2014; 82 (3-4): 474-479
- 29 Wanibuchi M, Fukushima T, Friedman AH. et al. Hearing preservation surgery for vestibular schwannomas via the retrosigmoid transmeatal approach: surgical tips. Neurosurg Rev 2014; 37 (03) 431-444 , discussion 444
- 30 Matsushima K, Kohno M, Nakajima N. Hearing preservation in vestibular schwannoma surgery via retrosigmoid transmeatal approach. Acta Neurochir (Wien) 2019; 161 (11) 2265-2269
- 31 Jia C, Xu C, Wang M, Chen J. How to precisely open the internal auditory canal for resection of vestibular schwannoma via the retrosigmoid approach. Front Surg 2022; 9: 889402
- 32 Seoane E, Rhoton Jr AL. Suprameatal extension of the retrosigmoid approach: microsurgical anatomy. Neurosurgery 1999; 44 (03) 553-560
- 33 Chanda A, Nanda A. Retrosigmoid intradural suprameatal approach: advantages and disadvantages from an anatomical perspective. Neurosurgery 2006; 59 (1, suppl 1): ONS1-ONS6 , discussion ONS1–ONS6
- 34 Samii M, Tatagiba M, Carvalho GA. Retrosigmoid intradural suprameatal approach to Meckel's cave and the middle fossa: surgical technique and outcome. J Neurosurg 2000; 92 (02) 235-241
- 35 Ishi Y, Terasaka S, Motegi H. Retrosigmoid intradural suprameatal approach for petroclival meningioma. J Neurol Surg B Skull Base 2019; 80 (Suppl. 03) S296-S297
- 36 Xu Y, Hendricks BK, Nunez MA, Mohyeldin A, Fernandez-Miranda JC, Cohen-Gadol AA. Microsurgical anatomy of the endoscopy-assisted retrosigmoid intradural suprameatal approach to the Meckel's cave. Oper Neurosurg (Hagerstown) 2021; 21 (02) 41-47
- 37 Samii M, Metwali H, Samii A, Gerganov V. Retrosigmoid intradural inframeatal approach: indications and technique. Neurosurgery 2013; 73 (1, suppl Operative): ons53-ons59 , discussion ons60
- 38 Colasanti R, Tailor AR, Gorjian M, Zhang J, Ammirati M. Microsurgical and endoscopic anatomy of the extended retrosigmoid inframeatal infratemporal approach. Neurosurgery 2015; 11 (Suppl. 02) 181-189 , discussion 189
- 39 Sato Y, Mizutani T, Shimizu K, Freund HJ, Samii M. Retrosigmoid intradural suprameatal-inframeatal approach for complete surgical removal of a giant recurrent vestibular schwannoma with severe petrous bone involvement: technical case report. World Neurosurg 2018; 110: 93-98
- 40 Meling TR, Zegarek G, Schaller K. How I do it: retrosigmoid intradural inframeatal petrosectomy. Acta Neurochir (Wien) 2021; 163 (03) 649-653
- 41 Matsushima K, Kohno M, Nakajima N. et al. Retrosigmoid intradural suprajugular approach to jugular foramen tumors with intraforaminal extension: surgical series of 19 cases. World Neurosurg 2019; 125: e984-e991
- 42 Constanzo F, Coelho Neto M, Nogueira GF, Ramina R. Microsurgical anatomy of the jugular foramen applied to surgery of glomus jugulare via craniocervical approach. Front Surg 2020; 7: 27
- 43 Jun W, Gao YL, Yu HG. et al. Comparison of translabyrinthine and retrosigmoid approach for treating vestibular schwannoma: a meta-analysis. Clin Neurol Neurosurg 2020; 196: 105994
- 44 Kim KH, Cho YS, Seol HJ. et al. Comparison between retrosigmoid and translabyrinthine approaches for large vestibular schwannoma: focus on cerebellar injury and morbidities. Neurosurg Rev 2021; 44 (01) 351-361
- 45 Irving RM, Jackler RK, Pitts LH. Hearing preservation in patients undergoing vestibular schwannoma surgery: comparison of middle fossa and retrosigmoid approaches. J Neurosurg 1998; 88 (05) 840-845
- 46 Ammirati M, Ma J, Cheatham ML, Maxwell D, Bloch J, Becker DP. Drilling the posterior wall of the petrous pyramid: a microneurosurgical anatomical study. J Neurosurg 1993; 78 (03) 452-455
- 47 Pillai P, Sammet S, Ammirati M. Image-guided, endoscopic-assisted drilling and exposure of the whole length of the internal auditory canal and its fundus with preservation of the integrity of the labyrinth using a retrosigmoid approach: a laboratory investigation. Neurosurgery 2009; 65 (06) 53-59 , discussion 59
- 48 Matsushima K, Komune N, Matsuo S, Kohno M. Microsurgical and endoscopic anatomy for intradural temporal bone drilling and applications of the electromagnetic navigation system: various extensions of the retrosigmoid approach. World Neurosurg 2017; 103: 620-630
- 49 Bernardo A, Boeris D, Evins AI, Anichini G, Stieg PE. A combined dual-port endoscope-assisted pre- and retrosigmoid approach to the cerebellopontine angle: an extensive anatomo-surgical study. Neurosurg Rev 2014; 37 (04) 597-608
- 50 Takemura Y, Inoue T, Morishita T, Rhoton Jr AL. Comparison of microscopic and endoscopic approaches to the cerebellopontine angle. World Neurosurg 2014; 82 (3-4): 427-441
Address for correspondence
Publication History
Article published online:
17 September 2024
© 2024. Asian Congress of Neurological Surgeons. This is an open access article published by Thieme under the terms of the Creative Commons Attribution-NonDerivative-NonCommercial License, permitting copying and reproduction so long as the original work is given appropriate credit. Contents may not be used for commercial purposes, or adapted, remixed, transformed or built upon. (https://creativecommons.org/licenses/by-nc-nd/4.0/)
Thieme Medical and Scientific Publishers Pvt. Ltd.
A-12, 2nd Floor, Sector 2, Noida-201301 UP, India
-
References
- 1 Basma J, Anagnostopoulos C, Tudose A. et al. History, variations, and extensions of the retrosigmoid approach: anatomical and literature review. J Neurol Surg B Skull Base 2021; 83 (Suppl. 02) e324-e335
- 2 Rodriguez Rubio R, Xie W, Vigo V. et al. Immersive surgical anatomy of the retrosigmoid approach. Cureus 2021; 13 (06) e16068
- 3 Al-Sharshahi ZF, Hoz SS, Alrawi MA, Sabah MA, Albanaa SA, Moscote-Salazar LR. The use of non-living animals as simulation models for cranial neurosurgical procedures: a literature review. Chin Neurosurg J 2020; 6: 24
- 4 Mantokoudis G, Huth ME, Weisstanner C. et al. Lamb temporal bone as a surgical training model of round window cochlear implant electrode insertion. Otol Neurotol 2016; 37 (01) 52-56
- 5 Cordero A, del mar Medina M, Alonso A, Labatut T. Stapedectomy in sheep: an animal model for surgical training. Otol Neurotol 2011; 32 (05) 742-747
- 6 Gocer C, Eryilmaz A, Genc U, Dagli M, Karabulut H, Iriz A. An alternative model for stapedectomy training in residency program: sheep cadaver ear. Eur Arch Otorhinolaryngol 2007; 264 (12) 1409-1412
- 7 Hamamcioglu MK, Hicdonmez T, Tiryaki M, Cobanoglu S. A laboratory training model in fresh cadaveric sheep brain for microneurosurgical dissection of cranial nerves in posterior fossa. Br J Neurosurg 2008; 22 (06) 769-771
- 8 Turan Suslu H, Ceylan D, Tatarlı N. et al. Laboratory training in the retrosigmoid approach using cadaveric silicone injected cow brain. Br J Neurosurg 2013; 27 (06) 812-814 Erratum in: Br J Neurosurg. 2014;28(6):823. Bahrı, Yasar [corrected to Bayrı, Yasar]
- 9 Sudhakara Rao M, Chandrasekhara Rao K, Raja Lakshmi C, Satish Chandra T, Murthy PSN. Suitable alternative for human cadaver temporal bone dissection: comparative micro ear anatomy of cattle, pig and sheep with human. Indian J Otolaryngol Head Neck Surg 2019; 71 (04) 422-429
- 10 Mallet C, Cornette R, Guadelli J. Morphometrical distinction between sheep (Ovis aries) and goat (Capra hircus) using the petrosal bone: application on French Protohistoric sites. Int J Osteoarchaeol 2019; 29 (04) 525-537
- 11 Irugu DV, Singh AC, Sikka K, Bhinyaram J, Sharma SC. Establishing a temporal bone laboratory in teaching institutes to train future otorhinolaryngologists and fundamentals of temporal bone laboratory: considerations and requirements. Indian J Otolaryngol Head Neck Surg 2016; 68 (04) 451-455
- 12 Mowry SE, Woodson E, Gubbels S, Carfrae M, Hansen MR. A simple assessment tool for evaluation of cadaveric temporal bone dissection. Laryngoscope 2018; 128 (02) 451-455
- 13 Frithioff A, Frendø M, Pedersen DB, Sørensen MS, Wuyts Andersen SA. 3D-printed models for temporal bone surgical training: a systematic review. Otolaryngol Head Neck Surg 2021; 165 (05) 617-625
- 14 Frithioff A, Frendø M, Weiss K. et al. Effect of 3D-printed models on cadaveric dissection in temporal bone training. OTO Open 2021; 5 (04) X211065012
- 15 Okada DM, de Sousa AM, Huertas RdeA, Suzuki FA. Surgical simulator for temporal bone dissection training. Braz J Otorhinolaryngol 2010; 76 (05) 575-578
- 16 Alvernia JE, Pradilla G, Mertens P, Lanzino G, Tamargo RJ. Latex injection of cadaver heads: technical note. Neurosurgery 2010; 67 (2, suppl operative): 362-367
- 17 Sanan A, Abdel Aziz KM, Janjua RM, van Loveren HR, Keller JT. Colored silicone injection for use in neurosurgical dissections: anatomic technical note. Neurosurgery 1999; 45 (05) 1267-1271 , discussion 1271–1274
- 18 Tatagiba M, Roser F, Schuhmann MU, Ebner FH. Vestibular schwannoma surgery via the retrosigmoid transmeatal approach. Acta Neurochir (Wien) 2014; 156 (02) 421-425 , discussion 425
- 19 Matsushima K, Kohno M, Komune N, Miki K, Matsushima T, Rhoton Jr AL. Suprajugular extension of the retrosigmoid approach: microsurgical anatomy. J Neurosurg 2014; 121 (02) 397-407
- 20 Alimohamadi M, Samii M. Suprajugular extension of the retrosigmoid approach. J Neurosurg 2014; 121 (03) 764-765
- 21 Constanzo F, Gerhardt J, Ramina R. How I do it: retrosigmoid suprajugular approach to the jugular foramen. Acta Neurochir (Wien) 2019; 161 (11) 2271-2274
- 22 Colasanti R, Tailor AR, Zhang J, Ammirati M. Functional petrosectomy via a suboccipital retrosigmoid approach: guidelines and topography. World Neurosurg 2016; 87: 143-154
- 23 Day JD, Kellogg JX, Fukushima T, Giannotta SL. Microsurgical anatomy of the inner surface of the petrous bone: neuroradiological and morphometric analysis as an adjunct to the retrosigmoid transmeatal approach. Neurosurgery 1994; 34 (06) 1003-1008
- 24 Scerrati A, Lee JS, Zhang J, Ammirati M. Exposing the fundus of the internal acoustic meatus without entering the labyrinth using a retrosigmoid approach: is it possible?. World Neurosurg 2016; 91: 357-364
- 25 Scerrati A, Lee JS, Zhang J, Ammirati M. Microsurgical anatomy of the internal acoustic meatus as seen using the retrosigmoid approach. Otol Neurotol 2016; 37 (05) 568-573
- 26 Cueva RA, Chole RA. Maximizing exposure of the internal auditory canal via the retrosigmoid approach: an anatomical, radiological, and surgical study. Otol Neurotol 2018; 39 (07) 916-921
- 27 Miller RS, Pensak ML. An anatomic and radiologic evaluation of access to the lateral internal auditory canal via the retrosigmoid approach and description of an internal labyrinthectomy. Otol Neurotol 2006; 27 (05) 697-704
- 28 Savardekar A, Nagata T, Kiatsoontorn K. et al. Preservation of labyrinthine structures while drilling the posterior wall of the internal auditory canal in surgery of vestibular schwannomas via the retrosigmoid suboccipital approach. World Neurosurg 2014; 82 (3-4): 474-479
- 29 Wanibuchi M, Fukushima T, Friedman AH. et al. Hearing preservation surgery for vestibular schwannomas via the retrosigmoid transmeatal approach: surgical tips. Neurosurg Rev 2014; 37 (03) 431-444 , discussion 444
- 30 Matsushima K, Kohno M, Nakajima N. Hearing preservation in vestibular schwannoma surgery via retrosigmoid transmeatal approach. Acta Neurochir (Wien) 2019; 161 (11) 2265-2269
- 31 Jia C, Xu C, Wang M, Chen J. How to precisely open the internal auditory canal for resection of vestibular schwannoma via the retrosigmoid approach. Front Surg 2022; 9: 889402
- 32 Seoane E, Rhoton Jr AL. Suprameatal extension of the retrosigmoid approach: microsurgical anatomy. Neurosurgery 1999; 44 (03) 553-560
- 33 Chanda A, Nanda A. Retrosigmoid intradural suprameatal approach: advantages and disadvantages from an anatomical perspective. Neurosurgery 2006; 59 (1, suppl 1): ONS1-ONS6 , discussion ONS1–ONS6
- 34 Samii M, Tatagiba M, Carvalho GA. Retrosigmoid intradural suprameatal approach to Meckel's cave and the middle fossa: surgical technique and outcome. J Neurosurg 2000; 92 (02) 235-241
- 35 Ishi Y, Terasaka S, Motegi H. Retrosigmoid intradural suprameatal approach for petroclival meningioma. J Neurol Surg B Skull Base 2019; 80 (Suppl. 03) S296-S297
- 36 Xu Y, Hendricks BK, Nunez MA, Mohyeldin A, Fernandez-Miranda JC, Cohen-Gadol AA. Microsurgical anatomy of the endoscopy-assisted retrosigmoid intradural suprameatal approach to the Meckel's cave. Oper Neurosurg (Hagerstown) 2021; 21 (02) 41-47
- 37 Samii M, Metwali H, Samii A, Gerganov V. Retrosigmoid intradural inframeatal approach: indications and technique. Neurosurgery 2013; 73 (1, suppl Operative): ons53-ons59 , discussion ons60
- 38 Colasanti R, Tailor AR, Gorjian M, Zhang J, Ammirati M. Microsurgical and endoscopic anatomy of the extended retrosigmoid inframeatal infratemporal approach. Neurosurgery 2015; 11 (Suppl. 02) 181-189 , discussion 189
- 39 Sato Y, Mizutani T, Shimizu K, Freund HJ, Samii M. Retrosigmoid intradural suprameatal-inframeatal approach for complete surgical removal of a giant recurrent vestibular schwannoma with severe petrous bone involvement: technical case report. World Neurosurg 2018; 110: 93-98
- 40 Meling TR, Zegarek G, Schaller K. How I do it: retrosigmoid intradural inframeatal petrosectomy. Acta Neurochir (Wien) 2021; 163 (03) 649-653
- 41 Matsushima K, Kohno M, Nakajima N. et al. Retrosigmoid intradural suprajugular approach to jugular foramen tumors with intraforaminal extension: surgical series of 19 cases. World Neurosurg 2019; 125: e984-e991
- 42 Constanzo F, Coelho Neto M, Nogueira GF, Ramina R. Microsurgical anatomy of the jugular foramen applied to surgery of glomus jugulare via craniocervical approach. Front Surg 2020; 7: 27
- 43 Jun W, Gao YL, Yu HG. et al. Comparison of translabyrinthine and retrosigmoid approach for treating vestibular schwannoma: a meta-analysis. Clin Neurol Neurosurg 2020; 196: 105994
- 44 Kim KH, Cho YS, Seol HJ. et al. Comparison between retrosigmoid and translabyrinthine approaches for large vestibular schwannoma: focus on cerebellar injury and morbidities. Neurosurg Rev 2021; 44 (01) 351-361
- 45 Irving RM, Jackler RK, Pitts LH. Hearing preservation in patients undergoing vestibular schwannoma surgery: comparison of middle fossa and retrosigmoid approaches. J Neurosurg 1998; 88 (05) 840-845
- 46 Ammirati M, Ma J, Cheatham ML, Maxwell D, Bloch J, Becker DP. Drilling the posterior wall of the petrous pyramid: a microneurosurgical anatomical study. J Neurosurg 1993; 78 (03) 452-455
- 47 Pillai P, Sammet S, Ammirati M. Image-guided, endoscopic-assisted drilling and exposure of the whole length of the internal auditory canal and its fundus with preservation of the integrity of the labyrinth using a retrosigmoid approach: a laboratory investigation. Neurosurgery 2009; 65 (06) 53-59 , discussion 59
- 48 Matsushima K, Komune N, Matsuo S, Kohno M. Microsurgical and endoscopic anatomy for intradural temporal bone drilling and applications of the electromagnetic navigation system: various extensions of the retrosigmoid approach. World Neurosurg 2017; 103: 620-630
- 49 Bernardo A, Boeris D, Evins AI, Anichini G, Stieg PE. A combined dual-port endoscope-assisted pre- and retrosigmoid approach to the cerebellopontine angle: an extensive anatomo-surgical study. Neurosurg Rev 2014; 37 (04) 597-608
- 50 Takemura Y, Inoue T, Morishita T, Rhoton Jr AL. Comparison of microscopic and endoscopic approaches to the cerebellopontine angle. World Neurosurg 2014; 82 (3-4): 427-441