

Subscribe to RSS
DOI: 10.1055/s-0034-1368615
Influence of Gut Microbiota-Derived Ellagitanninsʼ Metabolites Urolithins on Pro-Inflammatory Activities of Human Neutrophils
Authors
Correspondence
Publication History
received 05 November 2013
revised 17 May 2014
accepted 25 May 2014
Publication Date:
04 July 2014 (online)

Abstract
Ellagitannin-rich products exhibit beneficial influence in the case of inflammation-associated diseases. Urolithins, metabolites of ellagitannins produced by gut microbiota, in contrary to high molecular weight hydrophilic parental polyphenols, possess well established bioavailability. Because of the important role of neutrophils in progression of inflammation, the influence of urolithins on their pro-inflammatory functions was tested. Urolithin B at a concentration of 20 µM showed significant inhibition of interleukin 8 and extracellular matrix-degrading enzyme MMP-9 production. It was also significantly active in prevention of cytochalasin A/formyl-met-leu-phenylalanine-triggered selectin CD62L shedding. Urolithin C was the only active compound towards inhibition of elastase release from cytochalasin A/formyl-met-leu-phenylalanine-stimulated neutrophils with 39.0 ± 15.9 % inhibition at a concentration of 5 µM. Myeloperoxidase release was inhibited by urolithins A and C (at 20 µM by 46.7 ± 16.1 and 63.8 ± 8.6 %, respectively). Urolithin A was the most potent reactive oxygen species release inhibitor both in formyl-met-leu-phenylalanine and 4β-phorbol-12β-myristate-R13-acetate-stimulated neutrophils. At the concentration of 1 µM, it caused reactive oxygen species level decrease by 42.6 ± 26.6 and 53.7 ± 16.0 %, respectively. Urolithins can specifically modulate inflammatory functions of neutrophils, and thus could contribute to the beneficial health effects of ellagitannin-rich medicinal plant materials and food products.
Introduction
Ellagitannin-rich medicinal plant extracts and food products are currently commercialized and consumed due to their potential positive effects on diseases possessing inflammatory background. In inflammation-associated cardiovascular diseases, such products as nuts, pomegranate, blackberries, strawberries, and oak-aged red wine, were proven to exhibit beneficial effect in many human interventional studies [1], [2], [3], [4], [5], [6]. A recent study conducted on people with a high cardiovascular risk has clearly shown, that regular nut intake (walnuts, almonds, and hazelnuts) significantly reduced the incidence of major cardiovascular events connected with excessive inflammatory response [7].
Although many in vitro studies conducted for extracts and single compounds deal with the explanation of the mechanism of action, the extrapolation of their results on in vivo processes raises many difficulties due to the not well-established bioavailability of ellagitannins [1]. Ellagitannins are hydrophilic, high-molecular-weight dietary polyphenols containing hexahydroxydiphenoyl subunits. They are subsequently transformed by intestinal microbiota to dibenzo[b,d]pyran-6-one derivatives, urolithins. It was established on human model that ellagitannins from strawberries, raspberries, oak-aged wine, pomegranate juice, and nuts could be transformed by intestinal microbiota to urolithins, which are the main biomarkers detected in human plasma following the intake of ellagitannin-rich food products and medicinal plants. These metabolites in contrary to ellagitannins are lipophilic compounds which have a good bioavailability and may be present in plasma in the range of 0.5 to 18.6 µM [8], [9], [10]. Because of their catechol-like structure after absorption they are a potential substrate for catechol-O-methyl transferase (COMT) enzyme, which increases lipophilicity and may lead to biological activity alterations [8]. The gut microbiota metabolites of ellagitannins, urolithins, whose bioavailability is firmly confirmed, appear to be an attractive target for research of the factors contributing to ellagitanninsʼ positive effects on silencing the inflammation process. Products of subsequent dehydroxylation during microbial metabolism, urolithin C (UC), urolithin A (UA), and urolithin B (UB) (3, 2, 1 hydroxyl groups, respectively) ([Fig. 1]) were chosen to determine their influence on a series of pro-inflammatory functions of ex vivo stimulated human neutrophils. Additionally methylated forms: 8-methyl-O-urolithin A (MUA) and 8,9-dimethyl-O-urolithin C (MUC) ([Fig. 1]) as potential metabolites were examined to evaluate structure-activity relations of the initial compounds.

Neutrophils are known to play an important role in the development of many diseases with inflammatory background. Neutrophils are the first leucocytes which infiltrate inflammated tissue and are crucial contributors of the inflammatory process leading to its progression. Their elevated pro-inflammatory activity was especially well established in atherogenesis and other cardiovascular diseases [11]. They are important collaborators in the formation of murine atherosclerotic lesions. The activation of neutrophils is also a crucial step in progression of ischemia reperfusion injury. After the ischemic event, the cascade of pro-inflammatory signaling factors activates endothelium and induces neutrophil recruitment. Infiltrating neutrophils are a source of reactive oxygen species (ROS), proteinases and cytokines, which play a detrimental role during reperfusion [11], [12]. Released by stimulated neutrophils, interleukin 8 (IL-8) expresses proatherogenic properties by promotion of leukocyte arrest and migration together with promotion of proliferation and migration of smooth muscle and endothelial cells. The elevated levels of ROS production by neutrophils were demonstrated on patients with clinical conditions associated with high risk of developing atherosclerosis [11]. Neutrophil derived myeloperoxidase (MPO) was shown not only to mediate acute inflammatory responses, but also to catalyze reactions that consume vascular nitric oxide, resulting in impaired endothelial function [13]. High levels of neutrophil-derived MPO were found in atherosclerotic lesions [14], and this enzyme is also known to be a critical contributor for the development and progression of coronary artery disease in smokers [15]. Elastase is a proteolytic enzyme, whose significantly enhanced release was determined in patients with unstable angina pectoris and acute myocardial infarction [16]. Neutrophilsʼ adherence to the vascular endothelium is the first step and an essential event of neutrophils trans endothelial migration to inflammation site. Selectins are responsible for the rolling of neutrophils along the endothelium, while integrins determine their firm adhesion and migration. Stimulated neutrophils demonstrate downregulation of L-selectin CD62L and upregulation of integrin CD11b [17].
Because activated neutrophils play an important role in the development of pathologies connected with excessive inflammatory response, to explain the mechanism of the beneficial effect of consumed ellagitannins, the influence of urolithins on production and secretion of pro-inflammatory factors by ex vivo stimulated neutrophils is examined.
Results
All compounds at tested concentrations did not express cytotoxic effect towards neutrophils both in long and short-term tests (histograms presenting propidium iodide assay results are provided in Figs. 1S and 2S, Supporting Information).
The inflammatory signaling processes being a consequence of neutrophils recruitment triggered by TLR4 receptor agonist LPS were targeted. A statistically significant influence was observed for UB, which inhibited the production of main neutrophil cytokine IL-8 at the concentrations of 5 and 20 µM by 23.6 ± 5.2 and 31.6 ± 4.1 %, respectively. UA and UC significant inhibitory activity towards IL-8 expression was only observed at a concentration of 20 µM (24.8 ± 11.6 % and 19.4 ± 3.4 % of production inhibition, respectively) ([Fig. 2]). Statistically significant inhibition of production of extracellular matrix (ECM) degrading enzyme MMP-9 was also observed for UB ([Fig. 3]), with 42.0 ± 17.2 % and 49.6 ± 19.8 % of production inhibition at the concentrations of 5 and 20 µM, respectively. UA and UC inhibitory activities at 20 µM were 32.9 ± 25.1 % and 22.1 ± 7.0 % release inhibition, respectively. In both experiments, methylated derivatives (MUA and MUC) showed no significant activity. Curcumin, used as a positive control, inhibited at the concentration of 20 µM LPS-triggered production of IL-8 and MMP-9 by 73.3 ± 14.0 and 60.8 ± 1.8 %, respectively.


The rolling and firm adhesion of neutrophils is an important event in the progression of inflammatory state. When challenged with the pro-inflammatory factor, the L-selectin (CD62L) molecule, which is responsible for rolling, is shed in neutrophils, while integrin (CD11b), stored in granules and responsible for firm adhesion, becomes expressed on the surface [17]. The prevention of CD62L shedding was observed only in cells incubated with UB and MUA. The activity of these compounds was comparable to quercetin used as a positive control ([Fig. 4]). No influence on cytochalasin A/formyl-met-leu-phenylalanine (f-MLP)-triggered increased CD11b expression was observed for all tested compounds (Fig. 4S, Supporting Information).

Elastase is the neutrophil serine proteinase responsible for ECM degradation and is stored in azurophilic granules ready for immediate release upon stimulation. The most potent influence on elastase release from cytochalasin A/f-MLP-stimulated neutrophils was exhibited by UC, which at concentrations of 5 and 20 µM showed statistically significant and dose-dependent (p = 0.003 144) inhibition (39.0 ± 15.9 and 66.6 ± 7.0 %, respectively). The observed effect was compared with that of quercetin (at 20 µM 44.8 ± 18.8 % of inhibition), a known elastase inhibitor (positive control). The methylation of UC weakened but not completely abolished the anti-elastase activity (28.7 ± 16.3 % release inhibition at 20 µM; p = 0.000 470 vs. UC at concentration of 20 µM). Other compounds were inactive at the tested concentrations ([Fig. 5]).

MPO is another pro-inflammatory ready-to-release enzyme stored in neutrophilsʼ azurophilic granules, being responsible for production of HClO from H2O2 and Cl− during respiratory burst. The statistically significant inhibition of MPO release was observed for UA and UC (at 20 µM 46.7 ± 16.1 and 63.8 ± 8.6 %, respectively). UA was active even at the concentration of 1 µM (17.9 ± 7.9 % inhibition). The methylation of hydroxyl groups significantly decreased the effect, although at the concentration of 20 µM, the inhibition was still observed for MUA and MUC (31.2 ± 16.4 and 23.5 ± 19.3 %, respectively; p = 0.028 163 and p = 0.000 213 vs. UA and UC at concentration of 20 µM, respectively). The effects of UA and UC were compared with known MPO release inhibitor gallic acid, which at the concentration of 5 and 20 µM showed 44.0 ± 8.5 and 81.1 ± 7.6 % of inhibition, respectively ([Fig. 6]).

The inhibition of ROS release from stimulated neutrophils was conducted using two different models: one with stimulation achieved by bacterial derived peptide f-MLP, which leads to generation of superoxide anion (O2 • −), H2O2, and HClO, another by direct kinase C activator 4β-phorbol-12β-myristate-R13-acetate (PMA), which results in O2 • − and H2O2 release. In both models, the most active were UA and MUA showing, at a concentration of 20 µM, almost complete inhibition of ROS generation. UC was also active but prevented ROS generation significantly more effectively in f-MLP-luminol model than in PMA-lucygenin one (at a concentration of 5 µM, 67.2 ± 11.3 and 39.0 ± 4.7 % inhibition, respectively, p = 0.000 171; at a concentration of 20 µM, 88.2 ± 6.1 and 51.0 ± 9.2 % inhibition, respectively, p = 0.000 171). Interestingly, the methylation of UC (MUC) led to the significantly greater (at 20 µM 96.2 ± 2.7 % inhibition, p = 0.000 144) reduction of ROS generation in PMA stimulation model. For UA, UC, MUA, and MUC, the observed activities were statistically much stronger than those determined for ascorbic acid (p < 0.001) ([Fig. 7]).
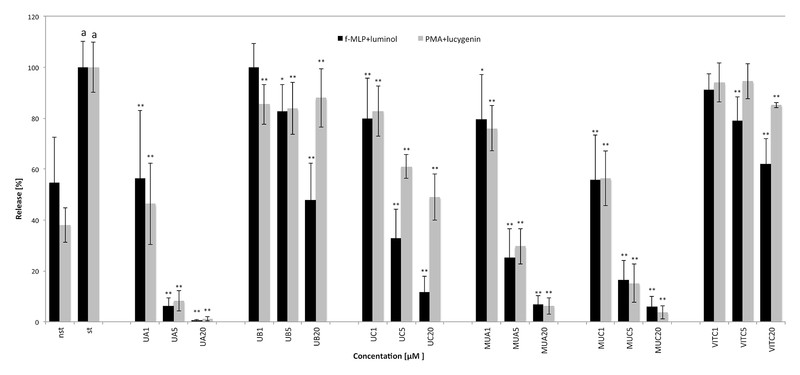
To determine whether the observed antioxidative effect on neutrophil model was triggered by inhibition of release or by direct scavenging activity, the influence of urolithins on in situ generated O2 • − by xanthin oxidase was examined. To exclude xanthin oxidase activity inhibition as a factor responsible for decrease of O2 • − levels, uric acid production was also determined. Significant scavenging activity was observed for all the compounds examined ([Fig. 8]). The SC50 values for UA, UB, UC, MUA, MUC, and ascorbic acid were 0.23 ± 0.11 µM, 0.72 ± 0.15 µM, 1.25 ± 0.68 µM, 0.77 ± 0.31 µM, 0.38 ± 0.21 µM, 6.68 ± 3.36 µM, respectively. For all compounds, the effect was dose-dependent and observed even at a concentration of 0.2 µM. Similarly as in neutrophils ROS generation model, the methylation of UA decreased the activity, while MUC was significantly more active than UC. Some effect on uric acid production was observed, but it was far too weak to be considered as responsible for such significant O2 • − levels decrease. The distribution of differences between compoundʼs scavenging effects was not entirely comparable to that observed in the neutrophil model but indicates that mostly direct scavenging is the mechanism of anti-ROS activity.

Discussion
The influence of urolithins on pro-inflammatory functions of neutrophils and observed variations of activity depending on compoundsʼ structures (number of substituents, hydroxyl groups, and methylation state) indicate the specificity of their pharmacological profiles. Urolithin B was the main metabolite responsible for inhibition of production of IL-8 and ECM degrading enzyme MMP-9. The compound was also active in prevention of CD62L shedding. Urolithin C was established as the only active elastase release inhibitor, and it expressed also significant inhibitory activity towards MPO release. It seems that in these models the presence of free hydroxyl groups is crucial for the observed effects, as the methylated compound was significantly less active. Moreover, according to previous observations made for flavonoids by Kanashiro et al. [18], elastase release inhibition by UC can be determined by its catechol group. It is interesting that in the case of anti-oxidant activity, the methylation did not alter the effect, or as in PMA-stimulated neutrophils model, the effect was significantly enhanced. UA was determined as the most potent anti-oxidant among the examined urolithins. The compound significantly inhibited release of ROS from neutrophils at a concentration of 1 µM in f-MLP and PMA stimulation models, and it was also a very potent MPO release inhibitor. In both experiments, activity depended on the presence of free hydroxyl groups, as MUA was less active than UA. On the other hand UA was shown to possess less significant impact on IL-8 and MMP-9 production than UB.
It needs to be noted that the complementary effect results from the influence on different neutrophil-associated pro-inflammatory functions, as all examined metabolites were previously detected in human plasma and are able to simultaneously interact with neutrophils in bloodstream.
In vivo experiments on animal model revealed strong activity of orally administered UA on reducing carrageenan-induced paw edema in mice [19]. UC and UB were shown to influence inflammation process on epigenetic level by inhibition of histone acetyltransferase activity [20]. In the study conducted on human aortic endothelial cells, UA was shown to inhibit TNFα-induced endothelial cell migration and to decrease the expression of chemokine (C–C motif) ligand 2 and interleukin-8. Its glucuronide inhibited monocyte adhesion and endothelial cell migration in a significant manner, however UB and its glucuronide were inactive [21]. Dellʼagli et al. [22] examined the influence of UA, MUA, and UB on MMP-9 production by THP-1 cells stimulated with TNFα. The results correspond with the ones obtained in the present study, as both UA and UB occurred to be active on contrary to significantly weaker MUA. Studies conducted by Gonzalez-Sarrias et al. on human colonic fibroblasts model suggest that inhibition of NF-κB translocation to nucleus by urolithins can be the mechanism responsible for their anti-inflammatory effects [23]. Cerda et al. [10] did not observe anti-oxidant properties of gut microbiota-derived ellagitannin metabolites as well as Ito et al., who conducted series of cell-free antioxidant assays including scavenging of xanthin oxidase derived O2 • −. In the cited study, UA and UC were completely inactive [24]. Although the presented results stand in contrast to these two previous reports, it was clearly shown that urolithins (especially A and C) were very active not only towards O2 • − direct scavenging, but also significantly inhibited neutrophil oxidative burst (both stronger than ascorbic acid). Our observations are consistent with Bialońska et al., who examined the influence of urolithins on intracellular ROS formation on HL-60 cell line and obtained corresponding results indicating that the antioxidant activity of urolithins is a consequence of not only the number of hydroxyl groups, but also lipophilicity of the molecule [25]. Considering the presented results, the number of hydroxyl groups is essential to the antioxidant activity, whereas other structural features (presence of methoxyl substituents) also significantly influence its grade.
The obtained results clearly show that urolithins are able to specifically modulate neutrophils functions which take part in the development and progression of inflammatory state. The effects were observed in the range of concentrations which are reachable in bloodstream after ingestion of ellagitannin-rich products. These results indicate that urolithins are at least partially responsible for the observed beneficial effects of orally administered ellagitannin-rich medicinal plant extracts and food products.
Materials and Methods
Chemicals
UA, UB, UC, MUA, and MUC were synthetized according to Bialonska et al. [25]. The identity was confirmed by NMR and MS spectra. Camptothecin (98 % purity), luminol, curcumin, PMA, f-MLP, ascorbic acid (reagent grade), NBT (nitrobluetetrazolium), xanthine, xanthine oxidase, SAAVNA (N-succinyl-alanine-alanine-valine-p-nitroanilide), LPS, cytochalasin A, TMB (3,3′,5,5′-tetramethylbenzidine) liquid substrate system, Triton X-100, Hanksʼ balanced salt solution (HSSB), and RPMI 1640 medium were purchased from Sigma-Aldrich GmbH. Lucigenin and quercetin (> 95 % purity) were purchased from Carl Roth. Gallic acid (> 96 % purity) was purchased from ChromaDex. Propidium iodide was purchased from BD Biosciences. All substances used were of > 95 % purity. PBS was purchased from Gibco.
Isolation of human neutrophils
Peripheral venous blood was taken from healthy human donors (20–35 years old) in the Warsaw Blood Donation Centre. Donors did not smoke or take any medication. They were clinically confirmed to be healthy, and routine laboratory tests showed values within the normal range. The study conformed to the principles of the Declaration of Helsinki. Neutrophils were isolated with a standard method of dextran sedimentation prior to hypotonic lysis of erythrocytes and to centrifugation in a Ficoll Hypaque gradient [26]. The purity of neutrophils preparation was > 97 %, and viability measured by tryptan blue exclusion was > 98 %. Neutrophils were then resuspended in appropriate medium to perform selected assay.
Cytotoxicity
Cytotoxicity of urolithins was determined by standard flow cytometric probe using propidium iodide (PI) staining to distinguish cells with diminished membrane integrity according to method previously described by Shinella et al. [27]. For short-term cytotoxicity determination, neutrophils (4 × 105/mL) were incubated in PBS with compounds at concentration of 1, 5, and 20 µM for 1.5 h. For long term cytotoxicity, neutrophils (2 × 105/mL) were cultured in a 24-well plate in RPMI 1640 medium with 10 % FBS, 10 mM HEPES, and 2 mM L-glutamine for 24 h at 37 °C with 5 % CO2 in presence of compounds at concentrations of 1, 5, and 20 µM. After 24 h, the neutrophils were harvested and centrifuged (1500 rpm; 10 min; 4 °C), washed once with cold PBS and resuspended in 400 µL of PBS. 5 µL of PI (50 µg/mL) solution was added to cell suspension. After 15 minutes of incubation at room temperature, cells were analyzed by BD FACSCalibur flow cytometer (BD Biosciences); 10 000 events were recorded per sample. Cells that displayed high permeability to PI were expressed as a percentage of PI(+) cells. Camptothecin at concentration of 10 µM was used as a positive control.
IL-8 and MMP-9 production
Neutrophils (2 × 105/mL) were cultured in a 24-well plate in RPMI 1640 medium with 10 % FBS, 10 mM HEPES, and 2 mM L-glutamine in the absence or presence of LPS (100 ng/mL) for 24 h at 37 °C with 5 % CO2 in the absence or presence of compounds at final concentrations of 1, 5, or 20 µM added to 1 mL of cell suspension 1 h before the stimuli. After 24 h the neutrophils were harvested and centrifuged (2000 rpm; 10 min; 4 °C). The amount of IL-8 or MMP-9 into cell supernatants was measured by enzyme-linked immunosorbent assay (ELISA) following the manufacturerʼs instructions (R&D Systems). According to Aggarwal et al. [28] and Antoine et al. [29], curcumin was used as a positive control. DMSO at final concentration 0.2 % was used as a non-stimulated control.
Expression of adhesion molecules CD62L and CD11b/CD18
Influence of urolithins on the level of adhesion molecules on neutrophils surface was determined using flow cytometric method. 500 µL of cell suspension (1 × 106) in PBS buffer was incubated with the compound for 30 min at 37 °C prior to 30 min stimulation with 10 µL of cytochalasin A (5 µg/mL) and f-MLP (0.1 µg/mL). Neutrophils were marked with monoclonal antibody against CD62L-(APC)-conjugate or CD11b-(PE)-conjugate (both from Becton Dickinson) and incubated for 30 min at 4 °C in the dark. The cells were analyzed by flow cytometry FACSCalibur (Becton Dickinson), and data from 20 000 events were recorded. According to Liu et al. [17], quercetin was used as a positive control. Three independent experiments were performed on neutrophils isolated from independent donors assayed in duplicate.
Elastase release
Neutrophil elastase release was determined using SAAVNA as a substrate, and p-nitrophenol was measured spectrophotometrically. 100 µL of cell suspension (5 × 105/mL) in HBSS was preincubated with 50 µL of compound solution (final concentrations of 1, 5, and 20 µM) for 15 min at 37 °C and then stimulated with 50 µL of cytochalasin A (5 µg/mL) and f-MLP (0.1 µg/mL) for 15 min. The neutrophils were centrifuged (2000 rpm; 10 min; 4 °C). After the addition of 50 µL of SAAVNA solution (1.6 mg/mL) to 100 µL of supernatant, the extent of p-nitrophenol was measured spectrophotometrically for 1 h at intervals of 20 min, at 412 nm using a microplate reader (BioTek). DMSO at final concentration 0.25 % was used as a non-stimulated control. According to Kanashiro et al. [18], quercetin was used as a positive control.
Myeloperoxidase release
Neutrophil MPO release was determined using TMB as a substrate. The assay is based on the oxidation of TMB by MPO in the presence of H2O2 [30]. 100 µL of cell suspension (2 × 105/mL) in HBSS was preincubated with 50 µL of compound solution (final concentrations of 1, 5, or 20 µM) for 15 min at 37 °C and then stimulated with 50 µL of cytochalasin A (5 µg/mL) and f-MLP (0.1 µg/mL) for 15 min. After centrifugation (2000 rpm; 10 min; 4 °C), 75 µL of supernatant was incubated with 100 µL of the TMB liquid substrate system. The reaction was terminated after 20 min by the addition of 2 M hydrochloric acid. The absorbance was measured at 655 nm using a microplate reader (BioTek). DMSO at final concentration 0.25 % was used as a non-stimulated control. According to Kroes et al. [31], gallic acid was used as a positive control.
Reactive oxygen species production
Oxidants generation by f-MLP or PMA-stimulated neutrophils was measured using luminol- or lucigenin-dependent chemiluminescence tests. 70 µL of cell suspension (2 × 105/mL) in HBSS was incubated with 50 µL of compound solution (final concentrations of 1, 5, and 20 µM) together with 50 µL of luminol (100 µM) or lucigenin (200 µM) solution. ROS production was initiated by the addition of 30 µL of f-MLP (0.1 µg/mL) or PMA (1 µg/mL). Changes in chemiluminescence at 37 °C were measured immediately for 45 min at intervals of 2 min in a microplate reader (Biotek). DMSO at final concentration 0.25 % was used as non-stimulated control. Ascorbic acid was used as a positive control.
Superoxide anion scavenging in cell-free systems
Scavenging of O2 • − was performed using a xanthine-xanthine oxidase system with the NBT reduction method as described by Choi et al. [32]. Compounds at concentrations of 0.05, 0.2, 1, 5, and 20 µM, 50 µL of xantine oxidase (0.04 U/ml in PBS), and 100 µL of 0.4 mM xantine and 0.24 mM NBT solution in PBS were added. The absorbance at 560 nm was measured in a microplate reader (BioTek) over a 30 min period. The percent of inhibition of the xantine/xantine oxidase system was calculated in comparison to the control without tested compounds. To evaluate whether compounds affected the O2 • − generation by direct interaction with xanthine oxidase, the enzyme activity was determined by monitoring the uric acid formation at 295 nm [33]. DMSO at final concentration 0.25 % was used as a control. Ascorbic acid was used as a positive control.
Statistical analysis
The results were presented as mean values ± SD. Statistical significance of differences between means was determined by one-way ANOVA with Tukeyʼs or Dunnettʼs post hoc tests. Results with p-value < 0.05 were considered statistically significant. All analyses were performed using Statistica 10 software.
Supporting information
Histograms presenting propidium iodide assay results for cytotoxicity (Figs. 1S and 2S) and data for non-active urolithins (Figs. 3S and 4S), as well as mean ± SD and p values (Tables 1S-6S) of urolithins effects corresponding to [Figs. 2]–[8] are available as Supporting Information.
Acknowledgments
The project was financially supported by Polish National Science Center research grant PRELUDIUM DEC-2011/03/N/NZ7/01785 and PhD scholarship ETIUDA decision number DEC-2013/08/T/ NZ7/00011.
Conflict of Interest
The authors declare that there are no conflicts of interest.
-
References
- 1 Larrosa M, Garcia-Conesa MT, Espin JC, Tomas-Barberan FA. Ellagitannins, ellagic acid and vascular health. Mol Aspects Med 2010; 31: 513-539
- 2 Porteri E, Rizzoni D, De Ciuceis C, Boari GE, Platto C, Pilu A, Miclini M, Agabiti Rosei C, Bulgari G, Agabiti Rosei E. Vasodilator effects of red wines in subcutaneous small resistance artery of patients with essential hypertension. Am J Hypertens 2010; 23: 373-378
- 3 Sabate J, Oda K, Ros E. Nut consumption and blood lipid levels: a pooled analysis of 25 intervention trials. Arch Intern Med 2010; 170: 821-827
- 4 Aviram M, Dornfeld L, Rosenblat M, Volkova N, Kaplan M, Coleman R, Hayek T, Presser D, Fuhrman B. Pomegranate juice consumption reduces oxidative stress, atherogenic modifications to LDL, and platelet aggregation: studies in humans and in atherosclerotic apolipoprotein E-deficient mice. Am J Clin Nutr 2000; 71: 1062-1076
- 5 Ma Y, Njike VY, Millet J, Dutta S, Doughty K, Treu JA, Katz DL. Effects of walnut consumption on endothelial function in type 2 diabetic subjects: a randomized controlled crossover trial. Diabetes Care 2010; 33: 227-232
- 6 Henning SM, Seeram NP, Zhang Y, Li L, Gao K, Lee RP, Wang DC, Zerlin A, Karp H, Thames G, Kotlerman J, Li Z, Heber D. Strawberry consumption is associated with increased antioxidant capacity in serum. J Med Food 2010; 13: 116-122
- 7 Estruch R, Ros E, Salas-Salvado J, Covas MI, Pharm D, Corella D, Aros F, Gomez-Gracia E, Ruiz-Gutierrez V, Fiol M, Lapetra J, Lamuela-Raventos RM, Serra-Majem L, Pinto X, Basora J, Munoz MA, Sorli JV, Martinez JA, Martinez-Gonzalez MA. Primary prevention of cardiovascular disease with a Mediterranean diet. N Engl J Med 2013; 368: 1279-1290
- 8 Seeram NP, Henning SM, Zhang YJ, Suchard M, Li ZP, Heber D. Pomegranate juice ellagitannin metabolites are present in human plasma and some persist in urine for up to 48 hours. J Nutr 2006; 136: 2481-2485
- 9 Tulipani S, Urpi-Sarda M, Garcia-Villalba R, Rabassa M, Lopez-Uriarte P, Bullo M, Jauregui O, Tomas-Barberan F, Salas-Salvado J, Espin JC, Andres-Lacueva C. Urolithins are the main urinary microbial-derived phenolic metabolites discriminating a moderate consumption of nuts in free-living subjects with diagnosed metabolic syndrome. J Agricult Food Chem 2012; 60: 8930-8940
- 10 Cerda B, Espin JC, Parra S, Martinez P, Tomas-Barberan FA. The potent in vitro antioxidant ellagitannins from pomegranate juice are metabolised into bioavailable but poor antioxidant hydroxy-6H-dibenzopyran-6-one derivatives by the colonic microflora of healthy humans. Eur J Nutr 2004; 43: 205-220
- 11 Baetta R, Corsini A. Role of polymorphonuclear neutrophils in atherosclerosis: Current state and future perspectives. Atherosclerosis 2010; 210: 1-13
- 12 Witko-Sarsat V, Rieu P, Descamps-Latscha B, Lesavre P, Halbwachs-Mecarelli L. Neutrophils: Molecules, functions and pathophysiological aspects. Lab Invest 2000; 80: 617-653
- 13 Eiserich JP, Baldus S, Brennan ML, Ma W, Zhang C, Tousson A, Castro L, Lusis AJ, Nauseef WM, White CR, Freeman BA. Myeloperoxidase, a leukocyte-derived vascular NO oxidase. Science 2002; 296: 2391-2394
- 14 van Leeuwen M, Gijbels MJ, Duijvestijn A, Smook M, van de Gaar MJ, Heeringa P, de Winther MP, Tervaert JW. Accumulation of myeloperoxidase-positive neutrophils in atherosclerotic lesions in LDLR−/− mice. Arterioscler Thromb Vasc Biol 2008; 28: 84-89
- 15 Rudolph TK, Rudolph V, Baldus S. Contribution of myeloperoxidase to smoking-dependent vascular inflammation. Proc Am Thorac Soc 2008; 5: 820-823
- 16 Dinerman JL, Mehta JL, Saldeen TG, Emerson S, Wallin R, Davda R, Davidson A. Increased neutrophil elastase release in unstable angina pectoris and acute myocardial infarction. J Am Coll Cardiol 1990; 15: 1559-1563
- 17 Liu JJ, Song CW, Yue Y, Duan CG, Yang J, He T, He YZ. Quercetin inhibits LPS-induced delay in spontaneous apoptosis and activation of neutrophils. Inflammation Res 2005; 54: 500-507
- 18 Kanashiro A, Souza JG, Kabeya LM, Azzolini AE, Lucisano-Valim YM. Elastase release by stimulated neutrophils inhibited by flavonoids: importance of the catechol group. Z Naturforsch C 2007; 62: 357-361
- 19 Ishimoto H, Shibata M, Myojin Y, Ito H, Sugimoto Y, Tai A, Hatano T. In vivo anti-inflammatory and antioxidant properties of ellagitannin metabolite urolithin A. Bioorg Medicinal Chem Lett 2011; 21: 5901-5904
- 20 Kiss AK, Granica S, Stolarczyk M, Melzig MF. Epigenetic modulation of mechanisms involved in inflammation: Influence of selected polyphenolic substances on histone acetylation state. Food Chem 2012; 131: 1015-1020
- 21 Gimenez-Bastida JA, Gonzalez-Sarrias A, Larrosa M, Tomas-Barberan F, Espin JC, Garcia-Conesa MT. Ellagitannin metabolites, urolithin A glucuronide and its aglycone urolithin A, ameliorate TNF-alpha-induced inflammation and associated molecular markers in human aortic endothelial cells. Mol Nutr Food Res 2012; 56: 784-796
- 22 Dellʼagli M, Galli GV, Bulgari M, Basilico N, Romeo S, Bhattacharya D, Taramelli D, Bosisio E. Ellagitannins of the fruit rind of pomegranate (Punica granatum) antagonize in vitro the host inflammatory response mechanisms involved in the onset of malaria. Malar J 2010; 9: 208
- 23 Gonzalez-Sarrias A, Larrosa M, Tomas-Barberan FA, Dolara P, Espin JC. NF-kappaB-dependent anti-inflammatory activity of urolithins, gut microbiota ellagic acid-derived metabolites, in human colonic fibroblasts. Brit J Nutr 2010; 104: 503-512
- 24 Ito H, Iguchi A, Hatano T. Identification of urinary and intestinal bacterial metabolites of ellagitannin geraniin in rats. J Agricult Food Chem 2008; 56: 393-400
- 25 Bialonska D, Kasimsetty SG, Khan SI, Ferreira D. Urolithins, intestinal microbial metabolites of Pomegranate ellagitannins, exhibit potent antioxidant activity in a cell-based assay. J Agricult Food Chem 2009; 57: 10181-10186
- 26 Boyum A. Isolation of mononuclear cells and granulocytes from human blood. Isolation of mononuclear cells by one centrifugation, and of granulocytes by combining centrifugation and sedimentation at 1 g. Scand J Clin Lab Invest 1968; 19: 125-131
- 27 Schinella G, Aquila S, Dade M, Giner R, Recio Mdel C, Spegazzini E, de Buschiazzo P, Tournier H, Rios JL. Anti-inflammatory and apoptotic activities of pomolic acid isolated from Cecropia pachystachya . Planta Med 2008; 74: 215-220
- 28 Aggarwal BB, Harikumar KB. Potential therapeutic effects of curcumin, the anti-inflammatory agent, against neurodegenerative, cardiovascular, pulmonary, metabolic, autoimmune and neoplastic diseases. Int J Biochem Cell Biol 2009; 41: 40-59
- 29 Antoine F, Simard JC, Girard D. Curcumin inhibits agent-induced human neutrophil functions in vitro and lipopolysaccharide-induced neutrophilic infiltration in vivo . Int Immunopharmacol 2013; 17: 1101-1107
- 30 Suzuki KO, Sasagawa S, Sakatani T, Fujikura T. Assay method for myeloperoxidase in human polymorphonuclear leukocytes. Anal Biochem 1983; 132: 345-352
- 31 Kroes BH, van den Berg AJ, Quarles van Ufford HC, van Dijk H, Labadie RP. Anti-inflammatory activity of gallic acid. Planta Med 1992; 58: 499-504
- 32 Choi CW, Kim SC, Hwang SS, Choi BK, Ahn HJ, Lee MY, Park SH, Kim SK. Antioxidant activity and free radical scavenging capacity between Korean medicinal plants and flavonoids by assay-guided comparison. Plant Sci 2002; 163: 1161-1168
- 33 Schepetkin IA, Kirpotina LN, Jakiw L, Khlebnikov AI, Blaskovich CL, Jutila MA, Quinn MT. Immunomodulatory activity of oenothein B isolated from Epilobium angustifolium . J Immunol 2009; 183: 6754-6766
Correspondence
-
References
- 1 Larrosa M, Garcia-Conesa MT, Espin JC, Tomas-Barberan FA. Ellagitannins, ellagic acid and vascular health. Mol Aspects Med 2010; 31: 513-539
- 2 Porteri E, Rizzoni D, De Ciuceis C, Boari GE, Platto C, Pilu A, Miclini M, Agabiti Rosei C, Bulgari G, Agabiti Rosei E. Vasodilator effects of red wines in subcutaneous small resistance artery of patients with essential hypertension. Am J Hypertens 2010; 23: 373-378
- 3 Sabate J, Oda K, Ros E. Nut consumption and blood lipid levels: a pooled analysis of 25 intervention trials. Arch Intern Med 2010; 170: 821-827
- 4 Aviram M, Dornfeld L, Rosenblat M, Volkova N, Kaplan M, Coleman R, Hayek T, Presser D, Fuhrman B. Pomegranate juice consumption reduces oxidative stress, atherogenic modifications to LDL, and platelet aggregation: studies in humans and in atherosclerotic apolipoprotein E-deficient mice. Am J Clin Nutr 2000; 71: 1062-1076
- 5 Ma Y, Njike VY, Millet J, Dutta S, Doughty K, Treu JA, Katz DL. Effects of walnut consumption on endothelial function in type 2 diabetic subjects: a randomized controlled crossover trial. Diabetes Care 2010; 33: 227-232
- 6 Henning SM, Seeram NP, Zhang Y, Li L, Gao K, Lee RP, Wang DC, Zerlin A, Karp H, Thames G, Kotlerman J, Li Z, Heber D. Strawberry consumption is associated with increased antioxidant capacity in serum. J Med Food 2010; 13: 116-122
- 7 Estruch R, Ros E, Salas-Salvado J, Covas MI, Pharm D, Corella D, Aros F, Gomez-Gracia E, Ruiz-Gutierrez V, Fiol M, Lapetra J, Lamuela-Raventos RM, Serra-Majem L, Pinto X, Basora J, Munoz MA, Sorli JV, Martinez JA, Martinez-Gonzalez MA. Primary prevention of cardiovascular disease with a Mediterranean diet. N Engl J Med 2013; 368: 1279-1290
- 8 Seeram NP, Henning SM, Zhang YJ, Suchard M, Li ZP, Heber D. Pomegranate juice ellagitannin metabolites are present in human plasma and some persist in urine for up to 48 hours. J Nutr 2006; 136: 2481-2485
- 9 Tulipani S, Urpi-Sarda M, Garcia-Villalba R, Rabassa M, Lopez-Uriarte P, Bullo M, Jauregui O, Tomas-Barberan F, Salas-Salvado J, Espin JC, Andres-Lacueva C. Urolithins are the main urinary microbial-derived phenolic metabolites discriminating a moderate consumption of nuts in free-living subjects with diagnosed metabolic syndrome. J Agricult Food Chem 2012; 60: 8930-8940
- 10 Cerda B, Espin JC, Parra S, Martinez P, Tomas-Barberan FA. The potent in vitro antioxidant ellagitannins from pomegranate juice are metabolised into bioavailable but poor antioxidant hydroxy-6H-dibenzopyran-6-one derivatives by the colonic microflora of healthy humans. Eur J Nutr 2004; 43: 205-220
- 11 Baetta R, Corsini A. Role of polymorphonuclear neutrophils in atherosclerosis: Current state and future perspectives. Atherosclerosis 2010; 210: 1-13
- 12 Witko-Sarsat V, Rieu P, Descamps-Latscha B, Lesavre P, Halbwachs-Mecarelli L. Neutrophils: Molecules, functions and pathophysiological aspects. Lab Invest 2000; 80: 617-653
- 13 Eiserich JP, Baldus S, Brennan ML, Ma W, Zhang C, Tousson A, Castro L, Lusis AJ, Nauseef WM, White CR, Freeman BA. Myeloperoxidase, a leukocyte-derived vascular NO oxidase. Science 2002; 296: 2391-2394
- 14 van Leeuwen M, Gijbels MJ, Duijvestijn A, Smook M, van de Gaar MJ, Heeringa P, de Winther MP, Tervaert JW. Accumulation of myeloperoxidase-positive neutrophils in atherosclerotic lesions in LDLR−/− mice. Arterioscler Thromb Vasc Biol 2008; 28: 84-89
- 15 Rudolph TK, Rudolph V, Baldus S. Contribution of myeloperoxidase to smoking-dependent vascular inflammation. Proc Am Thorac Soc 2008; 5: 820-823
- 16 Dinerman JL, Mehta JL, Saldeen TG, Emerson S, Wallin R, Davda R, Davidson A. Increased neutrophil elastase release in unstable angina pectoris and acute myocardial infarction. J Am Coll Cardiol 1990; 15: 1559-1563
- 17 Liu JJ, Song CW, Yue Y, Duan CG, Yang J, He T, He YZ. Quercetin inhibits LPS-induced delay in spontaneous apoptosis and activation of neutrophils. Inflammation Res 2005; 54: 500-507
- 18 Kanashiro A, Souza JG, Kabeya LM, Azzolini AE, Lucisano-Valim YM. Elastase release by stimulated neutrophils inhibited by flavonoids: importance of the catechol group. Z Naturforsch C 2007; 62: 357-361
- 19 Ishimoto H, Shibata M, Myojin Y, Ito H, Sugimoto Y, Tai A, Hatano T. In vivo anti-inflammatory and antioxidant properties of ellagitannin metabolite urolithin A. Bioorg Medicinal Chem Lett 2011; 21: 5901-5904
- 20 Kiss AK, Granica S, Stolarczyk M, Melzig MF. Epigenetic modulation of mechanisms involved in inflammation: Influence of selected polyphenolic substances on histone acetylation state. Food Chem 2012; 131: 1015-1020
- 21 Gimenez-Bastida JA, Gonzalez-Sarrias A, Larrosa M, Tomas-Barberan F, Espin JC, Garcia-Conesa MT. Ellagitannin metabolites, urolithin A glucuronide and its aglycone urolithin A, ameliorate TNF-alpha-induced inflammation and associated molecular markers in human aortic endothelial cells. Mol Nutr Food Res 2012; 56: 784-796
- 22 Dellʼagli M, Galli GV, Bulgari M, Basilico N, Romeo S, Bhattacharya D, Taramelli D, Bosisio E. Ellagitannins of the fruit rind of pomegranate (Punica granatum) antagonize in vitro the host inflammatory response mechanisms involved in the onset of malaria. Malar J 2010; 9: 208
- 23 Gonzalez-Sarrias A, Larrosa M, Tomas-Barberan FA, Dolara P, Espin JC. NF-kappaB-dependent anti-inflammatory activity of urolithins, gut microbiota ellagic acid-derived metabolites, in human colonic fibroblasts. Brit J Nutr 2010; 104: 503-512
- 24 Ito H, Iguchi A, Hatano T. Identification of urinary and intestinal bacterial metabolites of ellagitannin geraniin in rats. J Agricult Food Chem 2008; 56: 393-400
- 25 Bialonska D, Kasimsetty SG, Khan SI, Ferreira D. Urolithins, intestinal microbial metabolites of Pomegranate ellagitannins, exhibit potent antioxidant activity in a cell-based assay. J Agricult Food Chem 2009; 57: 10181-10186
- 26 Boyum A. Isolation of mononuclear cells and granulocytes from human blood. Isolation of mononuclear cells by one centrifugation, and of granulocytes by combining centrifugation and sedimentation at 1 g. Scand J Clin Lab Invest 1968; 19: 125-131
- 27 Schinella G, Aquila S, Dade M, Giner R, Recio Mdel C, Spegazzini E, de Buschiazzo P, Tournier H, Rios JL. Anti-inflammatory and apoptotic activities of pomolic acid isolated from Cecropia pachystachya . Planta Med 2008; 74: 215-220
- 28 Aggarwal BB, Harikumar KB. Potential therapeutic effects of curcumin, the anti-inflammatory agent, against neurodegenerative, cardiovascular, pulmonary, metabolic, autoimmune and neoplastic diseases. Int J Biochem Cell Biol 2009; 41: 40-59
- 29 Antoine F, Simard JC, Girard D. Curcumin inhibits agent-induced human neutrophil functions in vitro and lipopolysaccharide-induced neutrophilic infiltration in vivo . Int Immunopharmacol 2013; 17: 1101-1107
- 30 Suzuki KO, Sasagawa S, Sakatani T, Fujikura T. Assay method for myeloperoxidase in human polymorphonuclear leukocytes. Anal Biochem 1983; 132: 345-352
- 31 Kroes BH, van den Berg AJ, Quarles van Ufford HC, van Dijk H, Labadie RP. Anti-inflammatory activity of gallic acid. Planta Med 1992; 58: 499-504
- 32 Choi CW, Kim SC, Hwang SS, Choi BK, Ahn HJ, Lee MY, Park SH, Kim SK. Antioxidant activity and free radical scavenging capacity between Korean medicinal plants and flavonoids by assay-guided comparison. Plant Sci 2002; 163: 1161-1168
- 33 Schepetkin IA, Kirpotina LN, Jakiw L, Khlebnikov AI, Blaskovich CL, Jutila MA, Quinn MT. Immunomodulatory activity of oenothein B isolated from Epilobium angustifolium . J Immunol 2009; 183: 6754-6766