
Subscribe to RSS
DOI: 10.1055/s-0041-111631
Characterization of Neuraminidase Inhibitors in Korean Papaver rhoeas Bee Pollen Contributing to Anti-Influenza Activities In Vitro
Correspondence
Publication History
received 16 October 2015
revised 16 October 2015
accepted 23 November 2015
Publication Date:
05 February 2016 (online)
Abstract
The active constituents of Korean Papaver rhoeas bee pollen conferring neuraminidase inhibitory activities (H1N1, H3N2, and H5N1) were investigated. Six flavonoids and one alkaloid were isolated and characterized by nuclear magnetic resonance and mass spectrometry data. These included kaempferol-3-sophoroside (1), kaempferol-3-neohesperidoside (2), kaempferol-3-sambubioside (3), kaempferol-3-glucoside (4), quercetin-3-sophoroside (5), luteolin (6), and chelianthifoline (7). All compounds showed neuraminidase inhibitory activities with IC50 values ranging from 10.7 to 151.1 µM. The most potent neuraminidase inhibitor was luteolin, which was the dominant content in the ethyl acetate fraction. All tested compounds displayed noncompetitive inhibition of H3N2 neuraminidase. Furthermore, compounds 1–7 all reduced the severity of virally induced cytopathic effects as determined by the Madin-Darby canine kidney cell-based assay showing antiviral activity with IC50 values ranging from 10.7 to 33.4 µM (zanamivir: 58.3 µM). The active compounds were quantified by high-performance liquid chromatography, and the total amount of compounds 1–7 made up about 0.592 g/100 g bee pollen, contributing a rich resource of a natural antiviral product.
#
Key words
Papaver rhoeas - Papaveraceae - bee pollen - Apis mellifera - neuraminidase inhibitors - anti-influenza activity - flavonoid glycosidesAbbreviations
Introduction
Bee pollen is a granule-type material of agglutinated flower pollen made by worker honey bees with nectar and bee secretions. The resulting pollen is higher in nutritional value than untreated pollen, and is used as the primary source of protein for the bee hive. Bee pollen has been used in folk medicine from ancient times in many regions of the world for its medical properties to alleviate or cure conditions such as colds, flu, ulcers, and anemia [1]. Recently, biological effects such as antioxidation [2], anti-inflammation [3], antiallergy [4], antifungal [5], anticancer [6], chemopreventive [7], and antimutagenic [8] activities were also reported.
Like other apicultural products, royal jelly, honey, and propolis, the chemical composition of bee pollen depends on the plant source, regional vegetation, season, and honeybee races at the site of collection. It is rich in carbohydrates, proteins, lipids, minerals, vitamins, and various organic compounds, including phenolic acids and flavonoids. Most flavonoids from bee pollen exist as glycosides and contain kaempferol or quercetin as aglycone parts. Other phenolic acids usually found in floral parts, such as triacylated spermidine, were identified as major constituents of bee pollen [9].
Influenza viruses contain two surface antigens, NA and HA. NA and HA both recognize carbohydrate structures and bind to terminal sialic acid units on the surface of the host cell. When the influenza virus replicates, the binding of HA to its receptor initiates viral entry into the host cell, where NA catalyzes the cleavage of sialic acid. Since NA plays an important role in replication, spread, and pathogenesis during infection, blocking the function of NA is considered to be one of the most promising areas of targeting for the treatment of influenza. In the past decade, many reports have investigated substances related to NA inhibition. It was previously reported that flavonoids and (oligo) stilbenes are the most prominent natural products with NA inhibitory activity [10]. Therefore, pollen is considered one of the natural sources of NA inhibitors due to its high levels of polyphenols.
Papaver rhoeas L. is a species of flowering plant in the poppy family, Papaveraceae. It is sometimes mistaken for Papaver somniferum L., the opium poppy. It was reported that the flavonoids kaempferol, quercetin, luteolin, 8-hydroxyluteolin (hypolaetin), quercetin-3-glucoside (isoquercitrine), kaempferol-3-glucoside (astragaline), and quercetin-3-galactoside (hyperoside) were isolated from the methanol extract of the petals of Papaver rhoeas [11].
Although pollen has traditionally been used to treat the cold and flu, there are no reports of the antiviral activities of P. rhoeas bee pollen. In the present study, we investigated the active agents of Korean P. rhoeas bee pollen for inhibitory activity on NA from influenza virus A H1N1, H3N2, and H5N1, with an investigation of the anti-influenza viral activities in MDCK cells through a CPE reduction assay.
#
Results and Discussion
Influenza NA has been established as a primary drug target for the treatment of influenza infections. Cell culture-based assays need to be performed to evaluate the cytotoxicity and to confirm the anti-influenza activity of potential NAIs. Therefore, we first investigated the active constituents of Korean P. rhoeas bee pollen on NA inhibition. The neuraminidase inhibitors, compounds 1–7, from the ethyl acetate fraction of P. rhoeas bee pollen were isolated by chromatographic procedures, including purification via silica gel, Sephadex LH-20, ODS Sepak cartridges, and HPLC. The isolated compounds were identified as six flavonoids and one alkaloid by NMR and MS spectral data (see Supporting Information for details): kaempferol-3-sophoroside (kaempferol-3-O-β-D-glucosyl(1 → 2)-β-D-glucoside) (1), kaempferol-3-neohesperidoside (kaempferol-3-O-β-D-rhamnosyl(1 → 2)-β-D-glucoside) (2), kaempferol-3-sambubioside (kaempferol-3-O-β-D-xylosyl(1 → 2)-β-D-glucoside) (3), kaempferol-3-O-β-D-glucoside (4), quercetin-3-sophoroside (quercetin-3-O-β-D-glucosyl(1 → 2)-β-D-glucoside) (5), luteolin (6), and chelianthifoline (7), as shown in [Fig. 1]. Particularly, the glycoside part of the flavonoids isolated from P. rhoeas bee pollen was substituted on position 3 of the aglycone for kaempferol and quercetin. Compounds 1, 2, 3, and 5 had a common 1 → 2 linkage between biglycosides, which was established by HMBC spectral data. Compound 7 was previously reported as an active ingredient of Corydalis impatiens (Pall.) Fisch. ex DC. (Papaveraceae) [12] and as one of the urinary metabolites of protopine alkaloids unique to Papaver spp. [13].

The NA inhibitory activities of compounds 1–7 were evaluated against the recombinant influenza viral subtypes H1N1, H3N2, and H5N1, with zanamivir as a positive control. All compounds displayed inhibition of NAs in a dose-dependent manner. The IC50 values of compounds 1–7 on the NAs from influenza H1N1, H3N2, and H5N1 ranged from 10.7 to 100.5, 25.6 to 143.2, and 12.6 to 151.1 µM, respectively ([Table 1]). Compound 6, luteolin, exhibited the highest activity against the NAs of all three types. Comparison of the flavonoid glycosides (1–5) with the aglycone (6) indicated that the bulky sugar moiety in compounds 1–5 caused a loss of activity. However, compound 7 (chelianthifoline), an alkaloid, exhibited weak activity.
|
Compounds |
NA inhibition IC50 (µM) |
Inhibition type (ki, µM) |
Cytotoxicity and antiviral activity (µM) |
||||
|---|---|---|---|---|---|---|---|
|
H1N1 |
H5N1 |
H3N2 |
CC50 a |
IC50 b |
TIc |
||
|
Results were obtained from three independent experiments carried out in triplicate, and are expressed as the mean ± S. D.; a Concentration required to reduce cell growth by 50 %; b concentration required to inhibit virus-induced CPE by 50 %; c therapeutic index = CC50/IC50; d the compound used as the positive control; e not determined |
|||||||
|
1 |
85.6 ± 1.4 |
111.6 ± 1.0 |
61.0 ± 0.7 |
Noncompetitive (90.0 ± 1.9) |
> 100 |
21.3 ± 1.2 |
> 4.7 |
|
2 |
56.2 ± 1.1 |
80.2 ± 4.8 |
51.6 ± 1.5 |
Noncompetitive (57.8 ± 1.6) |
> 100 |
12.8 ± 4.2 |
> 7.8 |
|
3 |
45.3 ± 1.9 |
51.2 ± 3.1 |
52.2 ± 2.9 |
Noncompetitive (61.2 ± 0.7) |
> 100 |
33.4 ± 2.4 |
> 3.0 |
|
4 |
36.3 ± 1.5 |
61.1 ± 2.1 |
54.0 ± 1.2 |
Noncompetitive (56.7 ± 1.3) |
> 100 |
15.7 ± 4.2 |
> 6.4 |
|
5 |
88.3 ± 3.0 |
75.1 ± 8.7 |
112.8 ± 8.2 |
NDe |
> 100 |
12.5 ± 3.2 |
> 8.0 |
|
6 |
10.7 ± 0.2 |
12.6 ± 0.8 |
25.6 ± 8.3 |
Noncompetitive (46.1 ± 2.3) |
> 100 |
10.7 ± 1.6 |
> 9.4 |
|
7 |
100.5 ± 9.2 |
151.1 ± 19.8 |
143.2 ± 8.9 |
Noncompetitive (132.2 ± 0.7) |
> 100 |
21.5 ± 6.2 |
> 4.7 |
|
Zanamivird |
1.5 ± 0.2 nM |
3.4 ± 0.1 nM |
1.9 ± 0.4 nM |
ND |
> 100 |
58.3 ± 1.6 |
> 1.7 |
The manner of inhibition by the compounds was determined to evaluate the relative affinity for NA from the H3N2 influenza virus. To study the mode of inhibition, Dixon plots were used. As shown in [Fig. 2], all compounds tested (compounds 1–4, 6, and 7) displayed noncompetitive inhibitory activity, because the Vmax decreased while the Km remained the same with increasing concentrations of inhibitors, which intersected at a non-zero point on the x axis (Ki). The Ki values for these compounds were similar to their IC50 values ([Table 1]). To date, all flavonoids tested were reported to act noncompetitively. Recently, it was reported that essential functional groups of flavonoids to confer good NA inhibitory effects are hydroxyl groups in positions 4′ and 7, an oxo group in position 4, and a double bond between positions 2 and 3, showing agreement with our results [4].

The antiviral activities of compounds 1–7 against the influenza A virus were determined using the CPE reduction assay in MDCK cells to evaluate the cytotoxicity and to confirm the anti-influenza activity ([Table 1]). The IC50 values of these compounds were 10.7–33.4 µM, and the TI values were 2.99–9.35, which was 1.7- to 5.5-fold higher than that of zanamivir (IC50: 58.3 µM, TI: 1.7). Luteolin (6) was the most effective with an IC50 value of 10.7 µM (TI: > 9.35). Interestingly, alkaloid compound 7 demonstrated more potent activity in the CPE reduction assay than in the measurement of the neuraminidase inhibition effect. All of the compounds tested were not toxic to MDCK cells, displaying 100 % viability at the concentration of 100 µM ([Table 1]).
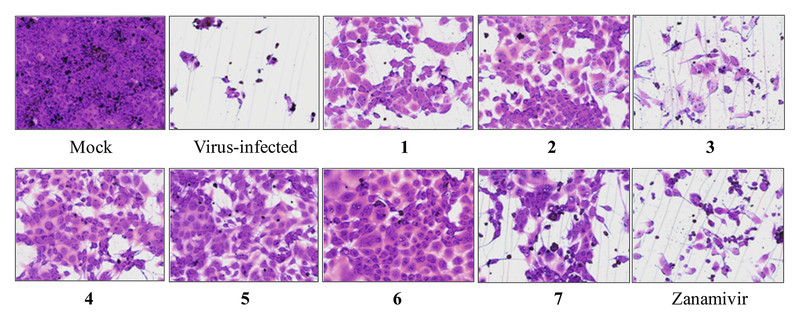
After 2 days of infection of MDCK cells with the influenza A virus, no differences were observed between the mock cells (non-virulent cells, [Fig. 3]) and those treated with 10 µM of the compounds (data not shown) in terms of normal morphology, with typical spread-out shapes. As shown in [Fig. 3], severe CPE was observed in influenza A virus-infected cells that were not treated with any of the compounds or with zanamivir. Compounds 2, 4, 5, and 6 potently inhibited the virus-induced CPE in infected MDCK cells ([Fig. 3]). Compounds 1 and 7 displayed moderate antiviral activities, while 3 only weakly reduced the CPE; however, their activities were comparable to that of zanamivir.
The qualification of the methanol extract and ethyl acetate fraction of bee pollen was performed by HPLC analysis on the basis of the retention time of the pure compounds and UV spectral data ([Fig. 4]). Compounds 1–3 and 6 (peaks 1–3 and 6) were dominant constituents in the ethyl acetate fraction, making up 55 % from the HPLC analysis data. Comparison of the two HPLC profiles revealed distinct differences in the peak of compound 6. Therefore, these results were consistent with the high neuraminidase inhibition shown by the ethyl acetate fraction of bee pollen. Peaks 8–10 were shown as main constituents (19 % from HPLC analysis of ethyl acetate fraction), but demonstrated no neuraminidase inhibitory activity up to 200 µM. The quantitative amount of compounds 1–7 found in the methanol extract of bee pollen is shown in [Table 2]. Calibration curves for compounds 1–7 were linear, as evidenced by the correlation coefficients (r2 = 0.997–0.999). The content of 1 was the highest (about 0.3 % of the total amount), while 2 and 5 were the major components, amounting to about 0.22 % of the total amount. Compounds 3, 6, and 7 were detected in small quantities, while 4 was not detected, as it was below 0.001 %. The total amount of active ingredients (1–7) of P. rhoeas bee pollen was about 0.592 g/100 g (dry bee pollen), which was considered to contribute as a rich resource of a natural antiviral product. The qualitative and quantitative analyses were carried out in triplicate with different batches and were reproducible.

|
Peak No. |
t R (min) |
Dried pollen (mg/100 g)a,b |
Identification |
|---|---|---|---|
|
a Values are means (n = 3); b standard deviation; c not detected; d not tested |
|||
|
1 |
16.8 |
294.9 ± 4.5 |
kaempferol-3-sophoroside (1) |
|
2 |
18.6 |
154.8 ± 1.3 |
kaempferol-3-neohesperidoside (2) |
|
3 |
20.1 |
24.5 ± 1.3 |
kaempferol-3-sambubioside (3) |
|
4 |
24.8 |
NDc (< 1) |
kaempferol-3-glucoside (4) |
|
5 |
12.9 |
63.2 ± 1.0 |
quercetin-3-sophoroside (5) |
|
6 |
32.7 |
24.9 ± 0.5 |
Luteolin (6) |
|
7 |
10.4 |
30.1 ± 0.9 |
Chelianthifoline (7) |
|
8 |
24.0 |
NTd |
Unknown |
|
9 |
27.0 |
NT |
Unknown |
|
10 |
29.6 |
NT |
Unknown |
Bee pollens have different chemical compositions depending on the plant source, regional vegetation, season, and honeybee races at the site of collection, and consequently, the biological activities of the bee pollens were different. Korean P. rhoeas bee pollen consisted of diverse flavonoid glycosides of high content exhibiting potent neuraminidase inhibition activity. Although these compounds were less active than zanamivir, a commercially available NA inhibitor, in our enzyme assay, they were more effective than zanamivir in the CPE reduction assay. This phenomenon suggests that the compounds isolated may have another mechanism of action in addition to neuraminidase inhibition. The detailed mechanism of action of these compounds still remains to be investigated. Importantly, this is the first report of the antiviral activity of P. rhoeas bee pollen, including NA inhibition and CPE reduction effects in influenza virus A-infected MDCK cells. Taken together, P. rhoeas bee pollen and its extract are good candidates for the development of novel prophylactic and therapeutic strategies against viral infection.
#
Materials and Methods
Pollen material
P. rhoeas bee pollen was provided and authenticated by emeritus professor Kwang-Ryul Choe at Chungnam National University, Daejeon, Korea, in 2011. P. rhoeas bee pollen was collected by honey bees, Apis mellifera L., at Chungnam province, Korea, in May-July 2011. A voucher specimen (Y-2011–01) was deposited at the herbarium of Chonbuk National University, Jeonbuk, Korea.
#
High-performance liquid chromatography analysis
The qualitative and quantitative analyses of the methanol extract of P. rhoeas bee pollen were characterized by the HPLC photo-diode array detection (PDA) system. For qualitative analysis, samples (10 µL) were injected into an analytical ODS column. The mobile phase was composed of 0.04 % trifluoro acetic acid in water (A) and methanol (B). The gradient conditions were as follows: 3 min, 20 % B; 35 min, 50 % B; 40 min, 50 % B, with a flow rate of 1 mL/min and the detection at 207 nm. For quantitative analysis, the standard curves of compounds 1–7 were prepared over a concentration range of 31.25–250 µg/mL with four different concentration levels. Samples (15 µL) were injected into an analytical ODS column. The mobile phases and detection conditions were as follows: 20 % aqueous methanol (0.04 % TFA) for 1, 5, and 7, 25 % aqueous methanol (0.04 % TFA) for 2 and 3, 30 % aqueous methanol (0.04 % TFA) for 4, and 40 % aqueous methanol (0.04 % TFA) for 6. The detection wavelengths were 286 nm for 7, 254 nm for 5, 264 nm for 1–4, and 349 nm for 6. A line for each compound was plotted using linear regression of the peak area vs. concentration. The coefficient of correlation (r2) was used to judge the linearity.
#
Extraction and isolation
The dried pollen (400 g) was extracted with 70 % methanol (1 L × 3) for 24 h at room temperature, and the solution was concentrated under reduced pressure to obtain a crude extract (150 g). The extract was partitioned successively with n-hexane, chloroform, ethyl acetate, and butanol. The ethyl acetate fraction exhibited the most potent inhibitory activity against H1N1 neuraminidase. Therefore, the ethyl acetate fraction (3 g) was then subjected to a C18 Sepak cartridge eluting with a gradient of increasing methanol (10–100 %) in water to give four fractions (Fr. I–IV) based on the silica TLC pattern and HPLC analysis. Fractions I and II were followed by reversed-phase preparative HPLC with 35 % aqueous MeOH (0.04 % TFA) and 40 % aqueous MeOH (0.04 % TFA), respectively, at a flow rate of 6 mL/min. Compounds 1 (t R 34 min, 33.2 mg), 2 (t R 44 min, 35.5 mg), 5 (t R 22 min, 13.7 mg), and 3 (t R 60 min, 26.1 mg) were isolated from fraction I, while 4 (t R 79 min, 22.3 mg) was obtained from fraction II. Fraction III was chromatographed on a Sephadex LH-20 column with MeOH, and then purified by reversed-phase preparative HPLC with a 50 ~ 80 % aqueous MeOH gradient (0.04 % TFA) at a flow rate of 8 mL/min to yield 7 (t R 26 min, 9 mg). Compound 6 (14.2 mg) was obtained by column chromatography of a silica gel eluted with CHCl3-MeOH (10 : 1) and a Sephadex LH-20 eluted with CHCl3:MeOH (1 : 1) from fraction IV.
#
Neuraminidase inhibitory activity
Neuraminidase inhibitory activity was estimated using a previously reported fluorometric (FL) method, with some modifications [14]. Recombinant influenza A virus H1N1, H3N2, and H5N1 NAs were purchased from R & D systems. The substrate for the FL assays, 4-methylumbelliferyl-N-acetyl-α-D-neuraminic acid (MUNANA), is cleaved by NA to release N-acetyl-α-D-neuraminic acid (NANA) and the fluorescent compound 4-methylumbelliferone (MU). Therefore, NA inhibition by an NA inhibitor causes a reduced fluorescence of the MU. The substrate, 50 µL of 0.2 mM MUNANA, was mixed with 90 µL of 50 mM Tris buffer (containing 200 mM NaCl, 5 mM CaCl2, pH 7.5) at room temperature. Ten microliters of the sample and 50 µL of H1N1 NA (50 ng/mL) were added to a well in a plate. The mixture was then recorded at the excitation and emission wavelengths of 365 nm and 445 nm, respectively, with a POLAR OPTIMA. In the case of H3N2 and H5N1 NAs, 25 mM MES buffer (containing 500 mM NaCl, 5 mM CaCl2, pH 6.5) and 50 mM MES buffer (containing 500 mM NaCl, 5 mM CaCl2, pH6.5) were used, respectively, and H5N1 required an activation period of 24 h at 37 °C before the assay. The other methods were the same as those for H1N1 NA. The 50 % inhibitory concentration (IC50) for enzymatic activity of neuraminidase was determined from the dose-response curve using Excel software.
Inhibition (%) = [1-(S-S0)/(C-C0)] × 100
Where C is the fluorescence of the control (enzyme and substrate) after 30 min of incubation, C0 is the fluorescence of the control at zero time, S is the fluorescence of the tested samples (enzyme, sample, and substrate) after incubation, and S0 is the fluorescence of the tested samples at zero time.
#
Virus and cell culture
The influenza A virus was provided by ATCC (American Type Culture Collection) and propagated in MDCK cells at 37 °C. MDCK cells were maintained in minimal essential medium (MEM) supplemented with 10 % fetal bovine serum (FBS) and 0.01 % antibiotic-antimycotic solution. Antibiotic-antimycotic solution, trypsin-EDTA, FBS, and MEM were purchased from Gibco BRL. Tissue culture plates were purchased from Falcon. SRB and zanamivir (purity of > 98 %) were purchased from Sigma-Aldrich.
#
Assays of antiviral activity and cytotoxicity
Antiviral activity and cytotoxicity were evaluated by the SRB method, which involves assessing the degree of reduction in the CPE [15]. Briefly, MDCK cells were seeded in 96-well culture plates at a concentration of 2 × 104 cells/well. The cells were then incubated for 18–24 h at 37 °C, after which the medium was removed and the cells were washed with PBS. After washing, 0.09 mL of diluted virus suspension and 0.01 mL of medium supplemented with trypsin-EDTA and compounds 1–7 or zanamivir, the positive control, were added in 10-fold dilutions ranging from 0.1 to 100 µM. Cells were incubated at 37 °C in 5 % CO2 for two days. After washing once with PBS, 100 µL of cold (− 20 °C) 70 % acetone were added to each well, and the cells were fixed for 30 min at − 20 °C. The acetone was then removed, and the 96-well plates were dried in an oven for 30 min. After drying, 100 µL of 0.4 % (w/v) SRB in 1 % acetic acid solution were added to each well, followed by incubation for 30 min at room temperature. Unbound SRB was then removed, and the plates were washed five times with 1 % acetic acid. Plates were then placed in a drying oven for 24 h. Bound SRB was solubilized with 100 µL of 10 mM unbuffered Tris-base solution, and the plates were incubated for 30 min. The resulting absorbance at 540 nm was obtained using a VERSAmax microplate reader, with the reference absorbance set to 620 nm. Zanamivir was used as a positive control in the CPE inhibition test.
The morphology was assessed using a light microscope at 20 × 10 magnification. Images were recorded with Leica Application Suite V2.7 software. The 50 % inhibitory concentration (IC50) and 50 % cytotoxic concentration (CC50) were calculated by regression analysis.
#
Supporting information
NMR assignments of compounds 1–7 are available as Supporting Information.
#
#
Acknowledgements
We thank emeritus professor Kwang-Ryul Choi (Chungnam National University) for support and identification of the bee pollen material. This work was supported by a grant from the Basic Research Program through the National Research Foundation of Korea (NRF), funded by the Ministry of Science, ICT & Future Planning (2012R1A1A3011559). This research was supported by “Research Base Construction Fund Support Program” funded by Chonbuk National University in 2015.
#
#
Conflict of Interest
The authors declare no conflict of interest.
* In-Kyoung Lee and Byung Soon Hwang contributed equally to this work.
-
References
- 1 Graikou K, Kapeta S, Aligiannis N, Sotiroudis G, Chondrogianni N, Gonos E, Chinou I. Chemical analysis of Greek pollen – antioxidant, antimicrobial and proteasome activation properties. Chem Cent J 2011; 5: 33-41
- 2 Morais M, Moreira L, Feás X, Estevinho LM. Honeybee-collected pollen from five Portuguese natural parks: palynological origin, phenolic content, antioxidant properties and antimicrobial activity. Food Chem Toxicol 2011; 49: 1096-1101
- 3 Maruyama H, Sakamoto T, Araki Y, Hara H. Anti-inflammatory effect of bee pollen ethanol extract from Cistus sp. of Spanish on carrageenan-induced rat hind paw edema. BMC Complement Altern Med 2010; 10: 30-41
- 4 Medeiros KC, Figueiredo CA, Figueredo TB, Freire KR, Santos FA, Alcantara-Neves NM, Silva TM, Piuvezam MR. Anti-allergic effect of bee pollen phenolic extract and myricetin in ovalbumin-sensitized mice. J Ethnopharmacol 2008; 119: 41-46
- 5 Ozcan M. Inhibition of Aspergillus parasiticus NRRL 2999 by pollen and propolis extracts. J Med Food 2004; 7: 114-116
- 6 Furusawa E, Chou SC, Hirazumi A, Melera A. Antitumour potential of pollen extract on lewis lung carcinoma implanted intraperitoneally in syngeneic mice. Phytother Res 1995; 9: 255-259
- 7 Pinto B, Cagiagli F, Riccio E, Reali D, Saric A, Balog T, Likic S, Scarpato R. Antiestrogenic and antigenotoxic activity of bee pollen from Cystus incanus and Salix alba as evaluated by the yeast estrogen screen and the micronucleus assay in human lymphocytes. Eur J Med Chem 2010; 45: 4122-4128
- 8 Pascoal A, Rodrigues S, Teixeira A, Feas X, Estevinho LM. Biological activities of commercial bee pollens: antimicrobial, antimutagenic, antioxidant and anti-inflammatory. Food Chem Toxicol 2014; 63: 233-239
- 9 Negri G, Teixeira EW, Alves ML, Moreti AC, Otsuk IP, Borguini RG, Salatino A. Hydroxycinnamic acid amide derivatives, phenolic compounds and antioxidant activities of extracts of pollen samples from Southeast Brazil. J Agric Food Chem 2011; 59: 5516-5522
- 10 Grienke U, Schmidtke M, Grafenstein SV, Kirchmair J, Liedl KR, Rollinger JM. Influenza neuraminidase: a druggable target for natural products. Nat Prod Rep 2012; 29: 11-36
- 11 Hillenbrand M, Zapp J, Becker H. Depsides from the petals of Papaver rhoeas . Planta Med 2004; 70: 380-382
- 12 Niu XF, Xu HB, Liu X, Fan T, Qi L. Isoquinoline alkaloids from Corydalis impatiens . Chem Nat Compd 2013; 49: 187-189
- 13 Wynne PM, Vine JH, Amiet RG. Protopine alkaloids in horse urine. J Chromatogr B Analyt Technol Biomed Life Sci 2004; 811: 85-91
- 14 Ryu YB, Kim JH, Park SJ, Chang JS, Rho MC, Bae KH, Park KH, Lee WS. Inhibition of neuraminidase activity by polyphenol compounds isolated from the roots of Glycyrrhiza uralensis . Bioorg Med Chem Lett 2010; 20: 971-974
- 15 Choi HJ, Song JH, Park KS, Kwon DH. Inhibitory effects of quercetin 3-rhamnoside on influenza A virus replication. Eur J Pharm Sci 2009; 37: 329-333
Correspondence
-
References
- 1 Graikou K, Kapeta S, Aligiannis N, Sotiroudis G, Chondrogianni N, Gonos E, Chinou I. Chemical analysis of Greek pollen – antioxidant, antimicrobial and proteasome activation properties. Chem Cent J 2011; 5: 33-41
- 2 Morais M, Moreira L, Feás X, Estevinho LM. Honeybee-collected pollen from five Portuguese natural parks: palynological origin, phenolic content, antioxidant properties and antimicrobial activity. Food Chem Toxicol 2011; 49: 1096-1101
- 3 Maruyama H, Sakamoto T, Araki Y, Hara H. Anti-inflammatory effect of bee pollen ethanol extract from Cistus sp. of Spanish on carrageenan-induced rat hind paw edema. BMC Complement Altern Med 2010; 10: 30-41
- 4 Medeiros KC, Figueiredo CA, Figueredo TB, Freire KR, Santos FA, Alcantara-Neves NM, Silva TM, Piuvezam MR. Anti-allergic effect of bee pollen phenolic extract and myricetin in ovalbumin-sensitized mice. J Ethnopharmacol 2008; 119: 41-46
- 5 Ozcan M. Inhibition of Aspergillus parasiticus NRRL 2999 by pollen and propolis extracts. J Med Food 2004; 7: 114-116
- 6 Furusawa E, Chou SC, Hirazumi A, Melera A. Antitumour potential of pollen extract on lewis lung carcinoma implanted intraperitoneally in syngeneic mice. Phytother Res 1995; 9: 255-259
- 7 Pinto B, Cagiagli F, Riccio E, Reali D, Saric A, Balog T, Likic S, Scarpato R. Antiestrogenic and antigenotoxic activity of bee pollen from Cystus incanus and Salix alba as evaluated by the yeast estrogen screen and the micronucleus assay in human lymphocytes. Eur J Med Chem 2010; 45: 4122-4128
- 8 Pascoal A, Rodrigues S, Teixeira A, Feas X, Estevinho LM. Biological activities of commercial bee pollens: antimicrobial, antimutagenic, antioxidant and anti-inflammatory. Food Chem Toxicol 2014; 63: 233-239
- 9 Negri G, Teixeira EW, Alves ML, Moreti AC, Otsuk IP, Borguini RG, Salatino A. Hydroxycinnamic acid amide derivatives, phenolic compounds and antioxidant activities of extracts of pollen samples from Southeast Brazil. J Agric Food Chem 2011; 59: 5516-5522
- 10 Grienke U, Schmidtke M, Grafenstein SV, Kirchmair J, Liedl KR, Rollinger JM. Influenza neuraminidase: a druggable target for natural products. Nat Prod Rep 2012; 29: 11-36
- 11 Hillenbrand M, Zapp J, Becker H. Depsides from the petals of Papaver rhoeas . Planta Med 2004; 70: 380-382
- 12 Niu XF, Xu HB, Liu X, Fan T, Qi L. Isoquinoline alkaloids from Corydalis impatiens . Chem Nat Compd 2013; 49: 187-189
- 13 Wynne PM, Vine JH, Amiet RG. Protopine alkaloids in horse urine. J Chromatogr B Analyt Technol Biomed Life Sci 2004; 811: 85-91
- 14 Ryu YB, Kim JH, Park SJ, Chang JS, Rho MC, Bae KH, Park KH, Lee WS. Inhibition of neuraminidase activity by polyphenol compounds isolated from the roots of Glycyrrhiza uralensis . Bioorg Med Chem Lett 2010; 20: 971-974
- 15 Choi HJ, Song JH, Park KS, Kwon DH. Inhibitory effects of quercetin 3-rhamnoside on influenza A virus replication. Eur J Pharm Sci 2009; 37: 329-333